| Zurück |
Impressum
Datenschutz
Typenkreise und biologischer Zeitcharakter
Wenig von dem Wissen über
die vorweltlichen Zeitalter der Erde ist
allgemeines Bildungsgut geworden. Nicht Viele von denen, die
sonst in Künsten und Wissenschaften wie im Leben Bescheid wissen,
haben auch eine fest umrissene Vorstellung von der Geschichte des
Lebens und der Erde, von den Umwandlungen und Umwälzungen, welche
die Oberfläche unseres Heimatsternes und seine Lebewesen im Lauf
der Jahrmillionen durchmachten mußten. Sie erstaunen fast,
wenn sie hören, wie klar in vielen Zügen sich das Bild
vorweltlicher Erd- und Lebensepochen schon abhebt von einer noch weit
älteren Erdurzeit, in die wir noch nicht hineinleuchten
vermögen. Sie erstaunen noch mehr, wenn sie hören,
daß die geschichtliche und die urgeschichtliche Menschenzeit,
soweit sie uns bisher überhaupt bis zu den roh zugehauenen
Steinsplittern des Eiszeitmenschen erschlossen wurde, vergleichsweise
doch nur ein letzter Augenblick in der Wandlung der Erdoberfläche
und des darüber gebreiteten Lebensteppichs ist. Sie haben
vielleicht auch durch allerlei populäre Bücher erfahren,
daß der Mensch sich aus niederen Tieren "entwickelte" und
daß die Geschlechter der Tiere und Pflanzen in vormenschlicher
Zeit in reicher Zahl und Mannigfaltigkeit schon die Erde
bevölkerten und auch fossil in den losen und festen
Gesteinsschichten der Erdrinde gefunden werden. Das alles hat in
ihrer Vorstellung aber doch mehr oder weniger den Charakter einer nicht
weiter in ihr Bildungsstreben eingreifenden Kuriosität, und sie
bleiben im allgemeinen weit davon entfernt, es ernsthaft durchzudenken,
sich einen plastischen Zeitbegriff an Hand urweltlicher Vergangenheit
zu schaffen und sich zu fragen, was eine solche Perspektive für
unsere ganze Daseinsauffassung bedeuten könnte.
Drei große Weltzeitalter (siehe
Tabelle) stehen heute dem Erdgeschichtsforscher deutlich
vor Augen. Aus ihnen weiß er zu berichten von einem nie
ruhenden Wechsel der Länder und Meere, von Gebirgsbildung und
Gebirgsabtragung, von Epochen erhöhter oder abgeschwächter
vulkanischer Tätigkeit, von periodischen Klimaausschlägen,
unter denen es bis an die Pole hinauf bald mild und warm, bald durch
das Eintreten von Eis- und Schneezeiten kühler war und
Gletschermassen sich auch über Länder schoben, die wir heute
in tropischer Wärme daliegen sehen. Die großen
Weltalter haben wieder ihre Einzelperioden, immer bezeichnet durch
unaufhaltsam sich ändernde Erdzustände und durch bestimmte
bald langlebige, bald kurzlebige Pflanzen- und Tiergeschlechter.

Fig.
1 (Bildquelle/-text:
"Urwelt, Sage und Menschheit"
von Edgar Dacqué, 8. Aufl. 1938, R. Oldenbourg)
Die zwischen den einzelnen Erdperioden gezogenen Striche bedeuten keine scharfen Grenzen im zeitlichen Ablauf der erdgeschichtlichen Zustände und Vorgänge, als deren Produkt die Formationen, also die gleichalten Gesteinsserien der Erde mit ihren pflanzlichen oder tierischen Fossileinschlüssen, verdeckt oder offen aus früheren Zeitaltern daliegen. Auch sind die einzelnen Zeitabschnitte und Weltalter unter sich zeitlich nicht gleich lang, wie es nach den regelmäßigen Rubriken auf der Tafel scheinen möchte. Wenn man das Känozoikum = 1 setzt, so darf man nach der durchschnittlichen Gesteinsmächtigkeit seiner Ablagerungen das Mesozoikum = 3,
das Paläozoikum = 12 nehmen. Eine absolute Zeitdauer, wonach der Nichtfachmann so gerne fragt, hat sich bisher noch nicht berechnen lassen.
Doch mag man schätzungsweise für das Quartär rund 500 000 Jahre, für das ganze Känozoikum (mit Einschluß des Quartärs)
5-8 Millionen Jahre rechnen; demnach für das Mesozoikum 15-24 Millionen und für das Paläozoikum 60-100 Millionen. Möglicherweise sind dies Minimalwerte, doch zeigen sie, bis auf welches geringe Maß die vielfach verbreiteten größeren Ziffern zurückzuführen sind. Die vorpaläozoischen großen Zeitalter bis zurück, zur Urzeit der Erde mit der hypothetischen ersten Krustenbildung um den glühenden Erdball übertreffen jene drei genannten Weltalter um ein Vielfaches an Zeitdauer. Aus ihnen kennt man jedoch wenig Sicheres und vor alleim keine klar definierbaren, Tier- und Pflanzenreste wie aus den drei ersteren Weltaltern, die man deshalb auch als geologisch-historische Zeit den früheren Ären des Eozoikums und Azoikums gegenüberstellt.
Die zwischen den einzelnen Erdperioden gezogenen Striche bedeuten keine scharfen Grenzen im zeitlichen Ablauf der erdgeschichtlichen Zustände und Vorgänge, als deren Produkt die Formationen, also die gleichalten Gesteinsserien der Erde mit ihren pflanzlichen oder tierischen Fossileinschlüssen, verdeckt oder offen aus früheren Zeitaltern daliegen. Auch sind die einzelnen Zeitabschnitte und Weltalter unter sich zeitlich nicht gleich lang, wie es nach den regelmäßigen Rubriken auf der Tafel scheinen möchte. Wenn man das Känozoikum = 1 setzt, so darf man nach der durchschnittlichen Gesteinsmächtigkeit seiner Ablagerungen das Mesozoikum = 3,
das Paläozoikum = 12 nehmen. Eine absolute Zeitdauer, wonach der Nichtfachmann so gerne fragt, hat sich bisher noch nicht berechnen lassen.
Doch mag man schätzungsweise für das Quartär rund 500 000 Jahre, für das ganze Känozoikum (mit Einschluß des Quartärs)
5-8 Millionen Jahre rechnen; demnach für das Mesozoikum 15-24 Millionen und für das Paläozoikum 60-100 Millionen. Möglicherweise sind dies Minimalwerte, doch zeigen sie, bis auf welches geringe Maß die vielfach verbreiteten größeren Ziffern zurückzuführen sind. Die vorpaläozoischen großen Zeitalter bis zurück, zur Urzeit der Erde mit der hypothetischen ersten Krustenbildung um den glühenden Erdball übertreffen jene drei genannten Weltalter um ein Vielfaches an Zeitdauer. Aus ihnen kennt man jedoch wenig Sicheres und vor alleim keine klar definierbaren, Tier- und Pflanzenreste wie aus den drei ersteren Weltaltern, die man deshalb auch als geologisch-historische Zeit den früheren Ären des Eozoikums und Azoikums gegenüberstellt.
Zahllos sind die Lebewesen, die
solcherweise in den Jahrmillionen
vorweltlicher Zeitalter über die ihr Gewand stets wechselnde Erde
dahingingen. Immer wieder neue Gestalten drängten sich
hervor, bald langsam, bald hastig dem Schoß der Erde
entquellend. Meeres- und Landtiere, Mollusken und Korallen,
Gewürm und Lurche, Vögel und Säugetiere sind uns in
fossilen Resten überliefert aus allen Zeiten - nur der Mensch
nicht; bloß in dürftigen Körper- und Werkzeugresten
ganz zuletzt, aus den spätesten Schichten, wie wenn sein Dasein
nur der letzte ausklingende Pulsschlag der lebenschaffenden Natur
wäre. Man beruft sich auf die Abstammungslehre und gibt
einen hypothetischen Stammbaum der Lebewesen, dessen Endglied der
Mensch sein soll - ein Spätgeborener.
Die von Linné im 18.
Jahrhundert geschaffene und später
nicht mehr grundlegend geänderte Einteilung der lebenden Formen in
Arten, Gattungen, Familien usw. wurde späterhin auch auf die
vorweltlichen, auf die fossilen Formen übertragen und hat in
dieses Wirrsal äußerlich einstweilen Ordnung gebracht.
Die hiermit aus dem Leben herausgehobenen abstrakten Systemgruppen
wurden im Laufe des 19. Jahrhunderts immer reicher vermehrt und durch
neu und neu hinzuströmendes fossiles Tier- und
Pflanzenmaterial. Die formale Abgegrenztheit und Starrheit der
Linnéschen Systemgruppen aber schien dahinzuschwinden, als die
Abstammungslehre alle lebendigen Formen als Glieder einer
zusammenhängenden Kette, als Äste und Zweige eines
natürlichen Stammbaumes aufzufassen suchte. Trotz der
hiermit scheinbar eingetretenen Verwischung fester Grenzen zwischen den
Arten, Gattungen und Familien brachte die Abstammungslehre doch nicht
etwa wieder die frühere Unübersichtlichkeit mit, sondern
lieferte nun Liniensysteme, an denen die vielen organischen Formen
geschichtlich, also entsprechend ihrem Auftreten in den Erdzeitaltern
aneinandergereiht wurden. Was erdgeschichtlich früher da
war, konnte nicht der Nachkomme der erdgeschichtlich Späteren
sein, und umgekehrt. So schien eine exakte Begründung des
Lebensstammbaumes gegeben, und man sollte denken, daß sich mit
dieser Methode alsbald klare, eindeutige Stammreihen ergeben
hätten.
Doch eine neue Verwirrung trat
ein. Man hatte zu einfach
gedacht. Denn man hielt die größere und geringere
Formähnlichkeit der Arten und Gattungen auch für den
unmittelbaren Ausdruck ihrer engeren oder weiteren
Blutsverwandtschaft. Reihte man aber die jetztweltlichen und die
vorweltlichen Arten, statt nach der Zeitfolge, nach ihrer
größeren und geringeren Formgleichheit aneinander, so
stimmte diese formale Reihenfolge nicht mehr oder nur in ganz seltenen
Fällen und auf ganz kurzen Linien mit der wahren geologischen
Zeitfolge überein. Auch ließen sich die gleichen Arten
zu ganz verschiedenen Formenreihen anordnen, je nach den
Körpermerkmalen, wonach man sie gerade genetisch zu beurteilen
versuchte. So mußte man etwa bei der vergleichenden
Betrachtung der Fußumwandlung in der Huftiergruppe eine fossile
Art als das Vorläuferstadium einer anderen ansehen; vergleicht man
aber statt des Fußes das Gebiß, so erschien hierin die in
der Fußentwicklung nachkommende Art nun ihrerseits wieder als ein
primitiveres Entwicklungsstadium. Man bezeichnet diese, stets
eine stammesgeschichtliche Verwirrung anrichtende Erscheinung als
Spezialisationskreuzung und hat nunmehr klar erkannt, daß in ihr
die prinzipielle Unmöglichkeit beschlossen liegt, aus der
äußerlichen Aneinanderreihung der Formbildung zu dem
theoretisch geforderten, aber auch erkenntniskritisch nicht haltbaren
echten Stammbaum der organischen
(1)
Typen zu gelangen. Zwar bekommt man durch die
Aneinanderfügung von Formstadien ideale Reihen, mit denen sich
abstrakt eine Formumwandlung klar veranschaulichen läßt,
aber sie erschließen uns nicht die wirklich naturhistorische
Herkunft einer Art oder Gattung aus der anderen, die lediglich in der
vorembryonalen "Keimbahn" verläuft. Die sichtbaren Formen
sind nur die Symbole hierfür. So lernte man, die Begriffe
Gleiches, Ähnliches und Formverwandtes von dem Begriff des
innerlich Verwandten trennen, welche, wie man sieht, in der
Körpergestalt durchaus nicht immer und gewiß nicht immer
unmittelbar ihren Ausdruck finden brauchen.
Wir wissen längst,
daß der Ursprung der Hauptäste und
vieler Nebenäste des Lebensreiches weit hinunter in immer dunkler
werdende Epochen der Vorwelt reicht; wir wissen auch längst,
daß vieles Neue unvermittelt, nicht mit Früheren
stammbaummäßig verknüpft, auftauchte. Aber man
macht sich immer noch nicht zu der rettenden Betrachtung frei, die uns
aus der Erfolglosigkeit aller Stammbaumkonstruktionen lösen kann:
die erdgeschichtlich gegebene Geschlechterfülle anzusehen als die
lebendige Auswirkung fest gegebener Grundtypen, die zwar während
der vorweltlichen Epochen in stets wechselnder Gestalt, jedoch ihr
Wesen stets bewahrend, frei nebeneinander standen und vermutlich nur in
einer unserem Forschen bisher noch nicht aufhellbar gewordenen
erdgeschichtlichen Urzeit, vor jenen drei großen Weltaltern,
genetisch verknüpft waren.
Stellt man sich entschieden auf
den Standpunkt einer solchen
Typentheorie, wie sie meines Erachtens die Paläontologie uns
aufnötigt - gleichgültig, ob man etwas Starres oder begrenzt
Flüssiges in den Typen sehen will - so könnte es scheinen,
als ob damit ein Rückschritt gegenüber der bisherigen
stammbaumdenkenden Lehre gemacht sei. Vielleicht wird damit auch
wirklich ein Schritt zurück von der bisherigen Anschauung gemacht
mit dem Gewinn, daß man von diesem wieder erreichten
ursprünglicheren, unbefangeneren, rückwärts liegenden
Betrachtungspunkt eine Aussicht gewinnt, welche durch die allzu
große Nähe der Deszendenzmauer bisher versperrt blieb.
Hält man daran fest, daß Typen von jeher nebeneinander
bestanden, wenigstens für die erdgeschichtlich sicher erkundeten
Zeiten; hält man weiter fest, daß die Typen, nachdem sie
einmal als organische Formen Fleisch und Blut angenommen hatten, sich
in immer neuen Gestalten zum Ausdruck brachten, ohne von da ab mit
anderen Typen genetisch verbunden zu sein; und endlich, daß sie
unter dem Bild einer Umwandlung immer wieder von Zeitalter zu Zeitalter
an andere Lebensverhältnisse angepaßt erschienen, bis sie
ausstarben, und daß nur insoweit die Evolutionstheorie gilt - so
leuchtet es zugleich auch ein, daß unter bestimmten Zeit- und
Lebensumständen die nebeneinander bestehenden Typenkreise in
konvergenter Weise ein gleichartiges Aussehen ihrer Gattungen,
gleichartige äußere Körpergestalt und oft gleichartige
Einzelorgane gewannen. Ob solche Formenkonvergenzen von den
äußeren Lebensumständen oder von einer inneren
gleichartigen konstitutiven Gestaltungskraft, von gleichen
Evolutionsstufen abhängen, ist hier für die Feststellung der
Tatsache zunächst belanglos. Ist ihnen aber in einem
bestimmten Zeitpunkt dasselbe Kleid, dasselbe habituelle Gebaren und
vielfach dasselbe mehr oder minder auffallende Einzelorgan zuteil
geworden, dann erscheinen viele oder alle Gattungen innerhalb solcher
Typenkreise so, als ob sie zu einer genetisch einheitlichen
Stammesgruppe gehörten, wie etwa Affen und Mensch, während
sie doch nur biologisch-habituell gleichartig sind oder sich sogar in
ihren Abkömmlingen überkreuzen können, unbeschadet ihrer
trotzdem weiterbestehenden evolutionistischen Wandlungsfähigkeit,
worin sie immer wieder ihre Grundkonstitution, ihren Typus, ihre
Entelechie manifestieren, einerlei, ob sie dabei formalähnlich
bleiben oder sich später in ihrer Gestalt wieder voneinander
entfernen.
Um eine klare Vorstellung von
dem Unterschied zwischen der
älteren, heute gewiß noch nicht überwundenen
stammesgeschichtlichen Auffassung einerseits und der Lehre von den
konvergierenden und wieder auseinandertretenden Typenkreisen
andererseits zu vermitteln, seien hier zwei Figuren gegeben.
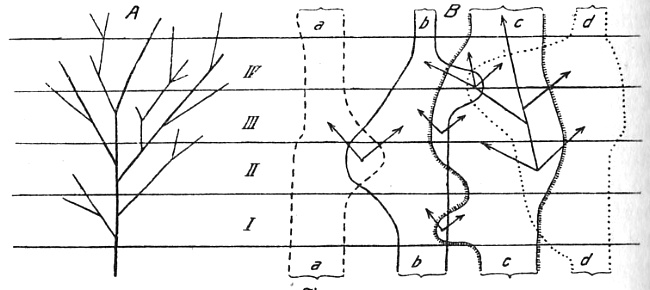
Fig. 2 (Bildquelle/-text: "Urwelt, Sage und
Menschheit"
von Edgar Dacqué, 8. Aufl. 1938, R. Oldenbourg)
Schema des Stammbaumes (A) und der sich überschneidenden Typenkreise (B) mit scheinbaren Stammreihen (Pfeile). I-IV geologische Zeiträume (s. obige Tabelle)
Schema des Stammbaumes (A) und der sich überschneidenden Typenkreise (B) mit scheinbaren Stammreihen (Pfeile). I-IV geologische Zeiträume (s. obige Tabelle)
Die eine (A)
liefert das Bild des Stammbaumes durch die
Zeitalter I-IV, und zwar so, als ob aus irgend einer konkreten Urform
sich der Lebensbaum entfaltet hätte. Die untersten Teile
wären früher niedere Tiere, nach oben folgten, aus ihnen
hervorgehend, immer höhere in immer größere
Mannigfaltigkeit; vielleicht zuletzt aus einem Primatenzweig der
Mensch. Wäre dieses Stammbaumbild als Ganzes oder in
Vielheit auf das Hervorkommen der organischen Formen im Lauf der
Erdgeschichte anwendbar, so müßten wir bei tieferem
Hinabsteigen in die Erdzeitalter immer weniger zahlreiche Formen
finden. Doch das Gegenteil ist der Fall: wir stoßen immer
wieder auf neue Typenkreise, die durchaus nicht stammbaumförmig
sich aneinanderreihen, wohl aber zu gleicher Zeit sich vielfach
gestaltlich in ihren Repräsentanten so begegnen, daß sie
sich formal verknüpfen lassen. Der Darstellung dieser
Erscheinung dient die andere Figurenhälfte (B): sie
veranschaulicht die typenhafte Selbständigkeit der den
natürlichen Stämmen zugrundeliegenden Entwicklungskreise a-d
und zeigt, in welcher Weise Übergangsformen, die man nach der
älteren Auffassung für stammesgeschichtliche
Abzweigungsstellen ansah, zustandekommen können. Es sind
einander formal überschneidende Evolutionen innerhalb jedes Typus,
nicht notwendig regelmäßig und in gleichem Umfang in
Erscheinung tretend, sondern unregelmäßig, von
äußeren Bedingungen vielleicht bestimmt und in
verschiedenerem Mengenumfang. Solche Formüberschneidungen
können auch an mehreren Stellen und in verschiedenen geologischen
Zeithöhen, vielleicht sogar wiederholt eintreten; es kommt dann
eine besonders verwirrende Fülle gleichartiger, aber ganz
verschiedenen Typenkreise (a-d) zugehöriger Gattungen zu gleicher
oder verschiedener geologischer Zeit zustande. Denkt man sich das
sphärisch und in seiner ganzen Plastik auf die Gestaltenbildung in
der Natur und auf die fossil vorliegenden Formen aus den Erdzeitaltern
übertragen, so bekommt man ein klares Bild davon, wie trotz fest
gebundener Lebenskreise, Lebenstypen, dennoch zu gleicher Zeit
außerordentlich ähnliche Gestalten, ohne unmittelbar
blutsverwandt zu sein und ohne einen unmittelbar zusammenhängenden
Stammbaum zu bilden, erscheinen können. Diese formalen
Überschneidungen sind es, welche immer und immer wieder zu den
prinzipiell verfehlten "Stammbäumen" Anlaß geben, wie sie
mittels der Pfeile angedeutet sind und die notwendigerweise
irreführen, weil sie keine wirklichen Entwicklungsbahnen
bezeichnen, sondern nur formale Ähnlichkeiten zusammenfassen und
die Umgrenzung der lebendig in sich geschlossenen Typenkreise nicht
sehen. Es seien Beispiele für solche
Formüberschneidungen gegeben oder, was dasselbe ist, für die
zu gleicher oder verschiedener Zeit immer wieder eintretende
ähnliche Organbildung oder Formgestaltung in heterogenen
Stammkreisen.
Im
paläozoischen Zeitalter, der ältestbekannten Epoche
vorweltlicher Lebensentwicklung, tritt bei verschiedenen, genetisch
nicht unmittelbar verbundenen Gruppen in der Schädelkapsel ein
Scheitelauge (Parietalorgan) auf.

Fig. 3 (Bildquelle/-text: "Urwelt, Sage und
Menschheit"
von Edgar Dacqué, 8. Aufl. 1938, R. Oldenbourg)
Panzer des Vorderkörpers eines paläozoischen Fisches mit zwei verschmolzenen Normalaugen; davor und dahinter je eine Stirn- und Scheitelöffnung.
Verkl. (Nach W. Patten. Mém. Acad. St. Pétersbg. 1903.)
Panzer des Vorderkörpers eines paläozoischen Fisches mit zwei verschmolzenen Normalaugen; davor und dahinter je eine Stirn- und Scheitelöffnung.
Verkl. (Nach W. Patten. Mém. Acad. St. Pétersbg. 1903.)
Zuerst erscheint
eine Stirnöffnung bei einigen
altpaläozoischen Fischen; bei anderen, die hierin wohl
ursprünglicher sind (Fig. 3), zugleich hinter den vereinigten
Normalaugen auch noch eine Scheitelöffnung; später bei den
Amphibien nur ein Scheitelloch, welche damit vollendet auf den Plan
treten. Sie behalten es bis in die Triaszeit hinein, wo es mehr
und mehr rudimentär wird. In der Permzeit kommen die
Reptilien hinzu, und diese besitzen es stets in voller Entwicklung
(Fig. 4), ebenfalls bis in die Triaszeit, wo es sich auch schon
häufig rückbildet.

Fig. 4 (Bildquelle/-text: "Urwelt, Sage und
Menschheit"
von Edgar Dacqué, 8. Aufl. 1938, R. Oldenbourg)
Fossiler Schädel eines Reptils der Permzeit (Casea) aus Texas mit besonders stark entwickeltem Scheitelloch. 2/3 nat. Gr.
(Nach S. W. Williston. Americ. Perm. Vertebrat. Chicago 1911.)
Fossiler Schädel eines Reptils der Permzeit (Casea) aus Texas mit besonders stark entwickeltem Scheitelloch. 2/3 nat. Gr.
(Nach S. W. Williston. Americ. Perm. Vertebrat. Chicago 1911.)
Alle Amphibien- und
Reptilformen nun, die man im mesozoischen Zeitalter
antrifft und die im Besitz eines vollentwickelten Organes sich halten,
sind wahrscheinlich Angehörige älterer, nämlich aus der
Permzeit schon stammender Typenkreise. So wäre also das
Spätpaläozoikum die bei vielen höheren Tieren das
Scheitelauge schaffende "Zeit". Es lebt aber heute noch auf
Neuseeland ein kleines, groteskes Reptil von einem Aussehen, wie wir es
den alten erdgeschichtlichen Reptilien mit ihrer Hautpanzerung und den
scharfen Konturen ihres Körpers und Kopfes vielfach beilegen
müssen. Diese Echse besitzt das Scheitelauge noch recht
deutlich, wenn auch schon in einem rudimentären Zustand
gegenüber den permisch-triassischen Formen. Aber sie ist
auch kein jungzeitliches Reptil. Sie gehört einem
Generaltypus an, der sich bis in das paläozoische Zeitalter hinein
zurückverfolgen läßt, dort sogar reichlich formbildend
war und auch das Mesozoikum in einigen Arten durchdauert.
Wüßten wir von ihrem Stamm aus früheren Zeiten nichts
und fänden wir erst heute diese Echse lebend, so könnten wir
allein aus dem Vorhandensein jenes so auffallenden und bisher
naturgeschichtlich immer noch nicht gedeuteten Organs alsbald Alter und
Herkunft ihres Typus angeben. In schwach rudimentärem
Zustand haben es auch die Eidechsen noch, und auch diese sind, wie die
Paläontologie anzunehmen Grund hat, aus gemeinsamer Wurzel mit
jenem Typus der neuseeländischen Echse zu paläozoischer Zeit
entstanden. Man sieht, welche bedeutsamen Ausblicke ein solches,
den Zeitcharakter vergleichend berücksichtigendes Verfahren bei
gehöriger künftiger Durcharbeitung bietet. Wenn aber
eine formbildende Epoche erst vorüber ist, so bekommt kaum je ein
später neu auftauchender Stamm ein solches Organ oder eine solche
Körpergestalt in derselben Weise wieder, wie es einer älteren
Epoche entsprach.
Es sei weiter auf
die Molchgestalt hingewiesen, die sich
äußerlich auszeichnet durch breit ausladende
Extremitäten nach Art des Salamanders. Die typische
Molchenzeit aber ist die letzte Hälfte des paläozoischen
Zeitalters. Da finden wir nicht nur "Molche" oder, besser gesagt,
Uramphibien mit den habituellen Merkmalen solcher, sondern zur selben
Zeit, zum Teil vergesellschaftet mit ihnen, echte Reptilien, welche
aber durch ihren breiten Kopf, ihre Körperhaltung, kurz durch ihre
ganze Tracht den Molch nachahmen, ohne zu seinem Stamm zu
gehören. Wieder eine andere Zeitepoche bringt die
Schildkrötengestalt hervor. Wie das
spätpaläozoische Zeitalter eine Molchgestalt, so schafft die
Triaszeit die Schildkröte. Denn nicht nur die echten, heute
noch als solche bezeichneten Schildkröten erscheinen damals
zuerst, um sich von da an weiter zu gestalten, sondern auch in einer
ganz anderen Gruppe kommen Schildkrötengestalten zum Vorschein,
die eben keine sind, sondern sich deren Kleid borgen. Oder wir
sehen seit der Alttertiärzeit in sehr verschiedenen
Säugetiergruppen, die man deshalb unter dem Namen Huftiere
zusammenfaßte, den fünf- und vierzehigen Fuß unter
Rückbildung seiner äußeren Zehen in den zweihufigen der
Rinder und Hirsche einerseits und in den einhufigen der Pferde
andererseits übergehen. Kein ein- oder zweizehiger
Unpaarhufer ist älter als die mitte der Tertiärzeit; und
obwohl jene Rückentwicklung auf den verschiedensten Stammlinien
unabhängig und parallel verlaufend sich vollzog, so blieben doch
die sich gleichenden Stadien an gleiche Zeitperioden gebunden und
erlauben daher bei Einzelfunden der Skelette eine sehr genaue
Bestimmung des geologischen Alter ihrer Lebenszeit. Die
mesozoische Epoche hinwiederum ist jenes Zeitalter der
Lebensentfaltung, wo unter den Landtieren der mehr oder minder
aufrechte Gang (Fig. 5) auf den beiden Hinterfüßen
angestrebt wird, im Gegensatz zu dem ursprünglich
gleichmäßigen Gang auf allen Vieren, wie ihn in primitivster
Weise der "Molchtypus" des spätpaläozoischen Zeitalters hatte.
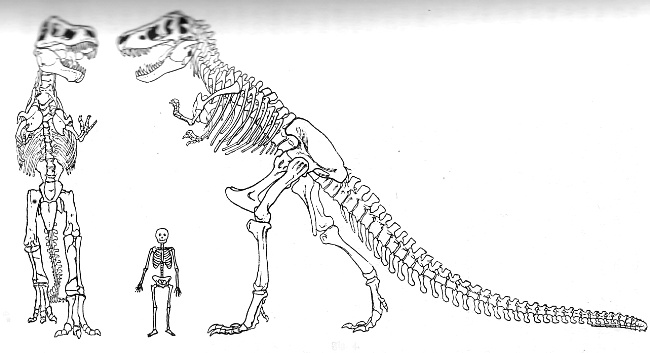
Fig. 5 (Bildquelle/-text: "Urwelt, Sage und
Menschheit"
von Edgar Dacqué, 8. Aufl. 1938, R. Oldenbourg)
Fossiles Reptilskelett von Raubtiercharakter (Tyrannosaurus) mit stark verlängertem Hinterbein bzw. sehr verkürzten Vorderbeinen und halb aufrechtem Gang.
Kreidezeit, Nordamerika. (Nach H. F. Osborn, Bull. Am. Mus. Nat. Hist. New York 1917.)
Zum Größenvergleich ist ein stark ausgewachsenes jetztweltliches Menschenskelett dazugestellt.
Fossiles Reptilskelett von Raubtiercharakter (Tyrannosaurus) mit stark verlängertem Hinterbein bzw. sehr verkürzten Vorderbeinen und halb aufrechtem Gang.
Kreidezeit, Nordamerika. (Nach H. F. Osborn, Bull. Am. Mus. Nat. Hist. New York 1917.)
Zum Größenvergleich ist ein stark ausgewachsenes jetztweltliches Menschenskelett dazugestellt.
Bei den Landtieren
des mesozoischen Zeitalters haben viele Gruppen
lange, kräftige Hinterbeine und kürzere Vorderbeine, wodurch
sie mit Unterstützung ihres kräftigen Schwanzes
aufrecht gehen konnten. Ja einige von ihnen haben hohle Knochen
wie Vögel, wobei schließlich in der Jurazeit auch
vogelähnliche Geschöpfe selbst erscheinen, vielleicht
gefiederte Reptilien mit langen Hinter- und kürzeren
Vorderextremitäten. Diese ganze Formengesellschaft
verrät also während des mesozoischen Zeitalters die Tendenz
zu jener Erhebung des Körpers auf den Hinterbeinen, wodurch das
mehr und mehr aufrechtgehende und das durch hohle Knochen erleichterte,
schließlich vogelähnliche Tier, mit zunehmender Befreiung
der Vorderextremität vom Boden, hervorgebracht wird.
Es gibt eine im
Lauf der Erdgeschichte nicht selten wiederkehrende
Erscheinung, die mit diesem Gesetz der Zeitcharakterbildung im Wesen
wohl gleichzusetzen ist: daß zu bestimmter Zeit eine gleichartige
Spezialform in verschiedenen Gruppen und Stämmen sich
herausbildet. Es ist gerade, als bedürfe die Natur an vielen
Stellen einer bestimmten Tiergestalt und präge sie aus irgend
welchen anderen Formen, die ihr gerade an den Plätzen zur
Verfügung stehen. Das bekannteste und auch anschaulichste
Beispiel ist die Nachahmung vieler höherer Säugetiertypen bei
der niedrigen Beuteltierfauna Australiens. Da finden wir einen
Beutelwolf, einen Beutellöwen, -bären, -dachse, -ratten,
-mäuse, fledermäuse, die alles das darstellen, um nicht zu
sagen nachahmen auf der Grundlage des Beuteltierkörpers, was jene
Tiergestalten des Wolfes, des Löwen, des Bären usw. in der
uns geläufigen höheren Säugetierwelt sind. "Das
Beuteltier als Wolf", "das Beuteltier als Ratte" - das wäre die
richtige Bezeichnung für diese eigentümlichen
Tierformen. Da solche nun schon in der Tertiärzeit da waren,
heute aber fast auf Australien beschränkt und bei uns sowie in
Amerika fast, in Asien ganz verschwunden sind, so hat sich daraus das
viel zitierte, aber auch viel mißverstandene Wort ergeben:
Australien stehe mit seiner Tierwelt noch im
Tertiärzeitalter. Natürlich ist dies irrig, weil die
Beuteltiere, anderwärts aussterbend, in Australien noch als
Relikten übrig blieben, aber nicht erst in der Quartärepoche
dort entstanden sind; denn nur die Entstehung solcher Formen ist
für die Tertiärzeit charakteristisch, und die gab es damals
auch schon in Australien; das Ausdauern kann in verschiedenen Gegenden
je nach den inneren und äußeren Lebensumständen der
Tiere verschieden lange währen.
Die Lemuriden, eine
gegenwärtig wesentlich auf Madagaskar
eingeschränkte Halbaffensippe, erscheint wie viele andere Typen
der Säugetiere, schon im frühesten Teil der
Tertiärzeit. Auch bei ihnen wird allerlei nachgeahmt.
Da entsteht das lemuroide Nagetier, das lemuroide Raubtier, die
lemuroide Fledermaus - alle mit entsprechendem Gebiß und
korrespondierenden Skelettmerkmalen, jedoch nie den Urtypus
verleugnend. Da die Signatur der Tertiärzeit bei den
Primaten, je länger je mehr, auch die Herausbildung des
anthropomorphen Affen ist und auch diese Evolution in
verschiedenstämmigen Gruppen konvergent vor sich geht, auch bald
mehr, bald weniger erreicht wird, so wird auch der Lemuridentyp von
solcher Formbildungstendenz ergriffen und stellt überdies noch
seine "Affenarten" heraus, die nun wie stammesgeschichtliche
Übergangsformen zu typenhaft echten Affen aussehen. Wie bei
den Lemuriden, so kommen auch in anderen Stammtypen während der
Tertiärzeit allerlei habituelle Charaktere dessen zum Vorschein,
was zuletzt die Menschenaffen repräsentieren und was auch der
Mensch in der Gestalt des Steinzeitmenschen teilweise noch an sich
trägt. Bald früher, bald später erscheint unter
der Säugetierwelt der Tertiärzeit in den Stammtypen das
Affenähnliche, und zwar umso mehr, je später es
geschieht. Alle sich an dieser Entwicklung ganz oder nur in
einzelnen Ästen beteiligenden Stämme und Stämmchen der
Säugetiere aber bleiben trotzdem als solche nebeneinander
bestehen, bilden je nach ihrer Grundkonstitution eigene Affenmerkmale
aus, wie auch der Menschenstamm selbst damals wohl am meisten
pithekoide Merkmale zur Schau trug und wahrscheinlich zur Entstehung
gewisser von da ab tierisch gebliebener Menschenaffen sein Teil
beitrug. So kamen jene gesuchten und teilweise gefundenen
formalen Übergangsglieder zustande, die man für
stammesgeschichtliche Bindeglieder zwischen Mensch und Affe ansah,
während es konvergente Formgestaltungen waren, im Sinne der Figur
2 B.
Wir haben also mit
dem Gesetz der Zeitformenbildung, das sich an fest
gegebenen Grundtypen unter dem Bild einer Entwicklung verwirklicht,
eine neue vergleichende Anatomie auch für den Menschen, die uns
sagen wird, wann er entstand. Wenn die bisherige Methode der
Betrachtung organischer Formen mehr und mehr das innerlich Typenhafte
vom äußerlich Konvergenten trennen lehrte, so wird durch die
neuartige Vergleichung der Formen nach ihrem Zeithabitus eine Art
biologischer Zeitsignatur für die einzelnen geologischen Epochen
festgestellt, die uns jeweils sichere Schlüsse auf das
Entstehungszeitalter eines Typus, eines Urformenstammes, ja eines
einzelnen Organes erlaubt, auch ohne daß wir durch Fossilfunde
selbst den gegenständlichen Nachweis für den
erdgeschichtlichen Augenblick des ersten Auftretens eines Typus
erbringen können. Wir sind somit auch imstande, aus
persistierenden oder rudimentären alten Form- und Organbildungen
der jetztzeitlichen Lebewesen, also auch des Menschen, das
Entstehungszeitalter des Stammes selbst zu ermitteln. Der
Zeitcharakter ist somit etwas durchaus Reales und nur einmal Gegebenes;
wer ihn späterhin noch trägt, ist seiner Zeitherkunft nach
daran erkennbar.
Ein weiteres, in
der Entwicklungslehre enthaltenes Problem ist der
Begriff des Höheren und Niederen, des Entwickelten und
Unentwickelten, Ausdrücke, die wie Selbstverständliches in
der Abstammungslehre gebraucht werden, jedoch ohne zureichende
erkenntniskritische Durchdringung. Auch diese beiden Begriffe
werden formalistisch und doch zugleich in einem absoluten Sinne
gebraucht. Man erklärt das amöboide Schleimtier
für niederer als das Molluskentier, dieses für niederer als
den Krebs, diesen für niederer als den Fisch, diesen für
niederer als den Lurch oder das Reptil, dieses für niederer als
das Säugetier und den Menschen. Das ist eine
unnaturwissenschaftliche Betrachtungsform gewesen, hinter der verkappt
mit dem Maßstab des Seelischen und des menschlich Geistigen oder
allenfalls nur des Ästhetischen gearbeitet wurde. Denn von
der Körperorganisation schlechthin aus gesehen ist es gar nicht
möglich, zu einer solchen absoluten Bewertung von Höherem und
Niederem zu kommen, und der nur naturwissenschaftliche mechanistische
Standpunkt gibt dafür überhaupt keinen Maßstab
her. Daß man auch in der nach mechanistischen Zielen
strebenden Deszendenzlehre den Menschen das höchstentwickelte
Geschöpf nannte, zeigt nur, wieviel metaphysisches Ausdeuten
gerade in dieser Lehre steckt, die von dem Begriff der "Entwicklung",
der durch und durch metaphysisch ist, trotz der formellen Verneinung,
niemals losgekommen ist. Ohne metaphysisch orientiertes
Werturteil ist kein Kriterium zu finden, wonach man ein Tier, einen
Organismus, der eben niemals Maschine ist, für höher oder
niederer erklären könnte. Das haben auch die
gänzlich metaphysiklosen Charakter unter den biologischen
Forschern der vergangenen Zeitepoche stets gefühlt, und deshalb
gab es auch in der hohen Zeit der dezendenztheoretischen Forschung
stets eine große Zahl von Gelehrten, die sogar eine
Stammbaumbeschreibung ablehnten und sich ausschließlich der
anatomisch vergleichenden Beschreibung der Arten widmeten, womit sie
die breite und sichere Grundlage schufen, von der aus allmählich
die formalistische, aber nicht biologische Abstammungslehre selbst
überwunden werden konnte.
Hiermit hängt
aufs engste auch die Klärung des Begriffes
Fortschritt zusammen, der gleichfalls einen naturhistorischen und einen
metaphysischen Sinn hat, die beide gerade in der Abstammungslehre
methodisch nicht klar auseinandergehalten worden sind. Solange
man die organische Welt als eine geschlossene Kette auseinander
hervorgehender Arten ansah, war das Linnésche System zugleich
das Idealbild des Stammbaumes. Als dieses ideale Bild unter den
Fossilfunden in Stammreihen zerfallen war, hielt man die Umwandlung
innerhalb dieser für einen Fortschritt und setzte den Begriff mit
dem der Entwicklung gleich. Zuletzt sah man, daß
Entwicklung und Fortschritt nur idealisierte Ausdrücke für
einseitige Spezialisation aufeinanderfolgender Formenstadien waren, und
nur so hat der Ausdruck Fortschritt derzeit überhaupt noch in der
Biologie einen greifbaren Wert. Fortschritt im streng
naturhistorisch-stammesgeschichtlichen Sinn ist aber nichts
Allgemeines, sondern stets eine Formbildung in bestimmter Richtung,
also geradezu eine Einseitigkeit; denn er geht immer auf Kosten und
unter Ausscheidung anderer Möglichkeiten, die oft biologisch nicht
weniger wichtig wären. Es wird bei einmal gegebenen Typen
von der Natur auf bestimmte Lebensformen und Anpassungen sozusagen
hingearbeitet, womit andere ausgeschlossen bleiben. Nicht anders
ist es ja auch im Leben der Völker und Kulturen: bestimmte
Grundanlagen werden entwickelt, das ist: spezialisiert. Latente
Möglichkeiten entfalten sich, andere bleiben unentwickelt oder
bleiben auf halbem Wege zurück oder verkümmern, je nach den
äußeren Bedingungen. So ist es in der organischen
Welt, und hierin ist kein Unterschied zwischen den Gattungen der
Lebewesen und den Lebensbildungen der Kulturen.
In diesem Sinne gibt es auch keine Rückwärtsentwicklung. Was aus der latenten Bindung im Typus entspringt und sich als Form gestaltet, kann sich halten oder wieder untergehen oder sich einseitig weiterbilden oder verkümmern; aber eine rückläufige Entwicklung des einmal Gestalteten, eine Rückkehr zum Ausgangspunkt und von da aus das Begehen eines neuen Entwicklungsweges, das gibt es in der Welt der organischen Bildungen niemals; sie sind alle geführt von dem unentrinnbaren Gesetz der Nichtumkehrbarkeit. Nur in der geistigen, in der sittlichen Welt gibt es die völlige Umkehr und das Verlassen des weit begangenen Weges unter großen Katastrophen oder inneren Erleuchtungen; aber in der Welt des Natürlich-Organischen kennen wir diese Freiheit nicht.
In diesem Sinne gibt es auch keine Rückwärtsentwicklung. Was aus der latenten Bindung im Typus entspringt und sich als Form gestaltet, kann sich halten oder wieder untergehen oder sich einseitig weiterbilden oder verkümmern; aber eine rückläufige Entwicklung des einmal Gestalteten, eine Rückkehr zum Ausgangspunkt und von da aus das Begehen eines neuen Entwicklungsweges, das gibt es in der Welt der organischen Bildungen niemals; sie sind alle geführt von dem unentrinnbaren Gesetz der Nichtumkehrbarkeit. Nur in der geistigen, in der sittlichen Welt gibt es die völlige Umkehr und das Verlassen des weit begangenen Weges unter großen Katastrophen oder inneren Erleuchtungen; aber in der Welt des Natürlich-Organischen kennen wir diese Freiheit nicht.
Wie aber
mußten unter solchen Umständen die Urformen
aussehen? Waren das neutrale Wesen, die noch nach keiner Richtung
spezialisiert waren, also - da jede nur denkbare Körperform an
sich schon eine Spezialisation ist - überhaupt keinen Körper
hatten?
Hier zeigt sich erst die begriffliche Unsicherheit der äußerlich verfahrenden alten Abstammungslehre. Denn ihr bedeutet Urform ein neutrales Geschöpf, aus dem durch fortgesetzte Zeugung und Umwandlung von Generation zu Generation schließlich spezialisierte Formen entstanden; der Streit dabei war nur der, ob diese Wandlung aus äußeren, also zufälligen, oder inneren konstitutionellen, nämlich im tieferen Sinn evolutionistischen Gründen geschehe. Wir aber verstehen unter Urform nicht einen solchen stammes-geschichtlich neutralen körperlichen Anfangspunkt, sondern die in allen zu einem Typus gehörigen Arten und Gattungen, auch in den anfänglichen, schon vollständig vorhandene typenhaft konstitutionelle Gebundenheit und Bestimmtheit, die Potenz, die bei allem äußeren evolutionistischen Formenwechsel als das Lebendig-Beständige das ist - eine Entelechie, wie auch Goethe wohl den Begriff Urform faßte. Es bekommt damit auch das deutsche Wort Entwicklung erst seinen tieferen, von der Sprache unbewußt schon erschlossenen Sinn zurück, als eine Manifestation des innerlich schon Vorhandenen.
Hier zeigt sich erst die begriffliche Unsicherheit der äußerlich verfahrenden alten Abstammungslehre. Denn ihr bedeutet Urform ein neutrales Geschöpf, aus dem durch fortgesetzte Zeugung und Umwandlung von Generation zu Generation schließlich spezialisierte Formen entstanden; der Streit dabei war nur der, ob diese Wandlung aus äußeren, also zufälligen, oder inneren konstitutionellen, nämlich im tieferen Sinn evolutionistischen Gründen geschehe. Wir aber verstehen unter Urform nicht einen solchen stammes-geschichtlich neutralen körperlichen Anfangspunkt, sondern die in allen zu einem Typus gehörigen Arten und Gattungen, auch in den anfänglichen, schon vollständig vorhandene typenhaft konstitutionelle Gebundenheit und Bestimmtheit, die Potenz, die bei allem äußeren evolutionistischen Formenwechsel als das Lebendig-Beständige das ist - eine Entelechie, wie auch Goethe wohl den Begriff Urform faßte. Es bekommt damit auch das deutsche Wort Entwicklung erst seinen tieferen, von der Sprache unbewußt schon erschlossenen Sinn zurück, als eine Manifestation des innerlich schon Vorhandenen.
Vom ersten
Augenblick ab, wo eine Urform in der lebenden Natur in
Gestalt einer oder mehrerer Arten sich darstellt, ist sie kein Schemen,
sondern ein Wesen mit Fleisch und Blut, voll Anpassung an die ihr
gemäßen Lebensumstände. Das Kennzeichen der
lebendigen Wesen ist gerade das, daß ihre natürliche
Körperhaftigkeit durch und durch biologisch zweckentsprechenden
Anpassungen und Spezialisierungen besteht, selbst wenn diese einen
geringen Grad der Vollendung gegenüber anderen, besser
angepaßten Formen haben oder sogar gelegentlich fehlgeschlagen
sind inbezug auf bestimmte Umweltbedingungen. Ein Typus wird
gewöhnlich von mehreren, unter verschiedenen Lebensbedingungen
stehenden Arten vertreten; doch selbst wenn er nur von einer einzigen
Art und wenigen Individuen vertreten wäre, so müßten
auch diese angepaßt und in ihrer Form nach irgend welchen
Richtungen spezialisiert sein, und wären sie auch die
allerältesten und primitivsten des Typus; denn anders können
und würden sie in der wirklichen Welt nicht gelebt haben.
Man darf daher nicht erwarten, irgendwo in der Erdgeschichte einmal
Urformen zu finden, die in ihrer Form neutral, nichtssagend,
schemenhaft gewesen wären. Stets hat sich das Leben in
wirklichen Charaktergestalten ausgelebt, die auch stets ihre eigene
Grundform variierten, ihren eigenen Typus verwirklichten, wenn er auch
noch so sehr durch biologische Zeitcharaktere verhüllt worden
wäre. Damit ist die stammes-geschichtliche Entwicklung nicht
verneint, sondern nur ihre unbiologische, allzu formalistische
Auffassung.
Das ist als
Hauptgesichtspunkt nun auch bei der Nachforschung nach
ältesten Menschengestalten obenan zu stellen. Auch diese
können selbst in ihrem denkbar ursprünglichsten Zustand
nichts weniger als Schemen gewesen sein, sondern müssen unbedingt
den menschlichen Typus, wenn auch unter mannigfachsten
Zeitgestaltungen, zum Ausdruck gebracht haben. Damit scheiden von
selbst alle Verknüpfungen des Urmenschen mit den Spättieren
aus, wobei man immer wieder das mit dem Menschen genetisch unmittelbar
nicht Zusammengehörige mit ihm verbinden, aus dem man ihn ableiten
will und womit man zu so grotesken Ergebnissen geführt wird, wie
dem, daß sein Typus irgendwie zu sehr später
erdgeschichtlicher Zeit aus spezialisierten Säugetieren
hervorgegangen sein könnte.
Es erschlossen sich
der bisherigen Abstammungslehre statt wirklicher
Stammbäume stets nur formal aneinandergereihte habituelle
Formenketten, während die persistenten Stammtypen als solche ihr
unsichtbar blieben, insbesondere der Menschentypus selbst, den die mehr
oder minder ähnlichen Affentypen am Ende der Tertiärzeit und
zuletzt wohl auch der nach der Affenseite hin degenerierte
Eiszeitmensch wie eine Folie umgeben, gewissermaßen die
Begleitung zur unterdessen schon unabhängig davon erklungenen
Melodie bildend. Der Menschenstamm tritt uns, fossil sichtbar,
spät erst in der Diluvialzeit entgegen, wohl weil er zuvor in
Gegenden lebte, die der geologischen Erforschung unzugänglich
geblieben sind. Wenn irgendwo, so hat uns gerade hier der
vielberufene Zufall des Findens und die notorische
Lückenhaftigkeit des Fossilmaterials einen Streich gespielt.
Gewöhnt man aber erst den Geist an die Unterscheidung von fest
gegebenen Grundtypen einerseits und biologischem Gewand andererseits,
das ihnen zeitweise und wechselnd übergeworfen ist, dann ist auch
die Bahn frei für eine richtigere Beurteilung der wahren Herkunft
des Menschen selbst und für die Feststellung, wie alt wohl sein
Stamm ist.
Das ergeschichtliche Alter des Menschenstammes
Fossile
Menschenreste, seien es Skeletteile oder nur Artefakte, sind,
wie schon erwähnt, bisher mit Sicherheit nur in den Ablagerungen
der Quartärzeit, dem letzten kurzen Abschnitt der Erdgeschichte,
nachgewiesen. Die letzte Phase dieser Quartärperiode ist die
Zeit der historischen Menschengeschichte, zurück bis zur
jüngeren Steinzeit. Diese selbst ist eine
prähistorische Epoche, aus der man geschliffene und polierte, also
gut ausgearbeitete Steinwerkzeuge hat. Dieser Periode voraus geht
das Diluvium mit einer Eiszeit als Hauptabschnitt der
Quartärepoche. Damals waren größere Flächen
der Nordhalbkugel und die meisten Hochgebirge bis in ihr Vorland hinaus
vergletschert; die durchschnittliche Jahrestemperatur war vermutlich
nur um 5° C kühler als jetzt. Diese Diluvialzeit mit
ihren wärmeren Zwischeneiszeiten ist nun die Periode der
altsteinzeitlichen Kulturen und fossiler Menschenrassen. Kurz
zuvor sollte sich, nach der gangbaren Lehre, der Mensch aus
spättertiärzeitlichen affenartigen Säugetieren
entwickelt haben; der Steinzeitmensch soll diesen Urformen noch
näher stehen.
Es wurde schon von
anderer Seite die Frage aufgerollt (2),
ob nicht die Dauer der
Steinzeitperiode bedeutend überschätzt werde und ob die
Entstehung der Kultur überhaupt erst mit dem Steinzeitalter
eingesetzt habe? Ist es möglich und denkbar, sagt Frobenius,
daß die aufkeimende Kultur bei einer Verbindung mit dem Stein,
also dem toten Teil des Erdkörpers begonnen hat? Liegt es
nicht viel näher, anzunehmen, daß das "Ur", der Anbeginn mit
einer Wechselbeziehung des menschlichen Könnens zu der lebendigen
Umwelt, zu Pflanze und Tier anhub? Und sollte es keine
Möglichkeit geben, das Umgekehrte der landläufigen Auffassung
aus dem Phänomen der Kultur abzulesen?
Freilich meint es Frobenius anders als wir; denn er hat noch nicht die hier erstrebte weite erdgeschichtliche Zeitperspektive. Er deutet nur allgemein an, daß die ältesten Werkzeuge nicht aus Stein, sondern eher aus tierischem und pflanzlichen Material erarbeitet worden sein könnten und daß daher die Steinzeit, auch die Altsteinzeit, ein verhältnismäßig spätes Entwicklungsstadium des "Urmenschen" gewesen wäre; denn zuerst und unmittelbar lebe der Mensch in dem feinen Schleier von Wasser, Erde, Pflanze und Tier, der, umspült vom Luftmeer und dünn, den harten Steinkörper der Erde umgibt. Diesem harten Körper stehe jener Schleier, zu dem der Mensch selbst gehört, als Einheit gegenüber, ausgesetzt dem Einfluß der Gestirne und von ihren Kräften bewegt. Wir Menschen sind, als Körper betrachtet, Ausdrucksform der Umbildungen dieses Schleiers; die Kultur aber ist der Ausdruck des in diesem Schleier bei seiner Beziehung zu einem "Jenseits" symbolisch sich auswirkenden Lebens. Demnach, so meint wohl Frobenius, hat die älteste, die erste Kulturregung zu ihrer Gestaltung unwillkürlich nach den Elementen und Teilen des Schleiers selbst gegriffen, nicht nach dem Material der nackten, vom Schleier entblößten Steinkugel. So erwartet er noch die Aufdeckung einer viel älteren vorsteinzeitlichen Kultur, und die müßte vor der Quartärepoche, also in der Tertiärzeit liegen.
Freilich meint es Frobenius anders als wir; denn er hat noch nicht die hier erstrebte weite erdgeschichtliche Zeitperspektive. Er deutet nur allgemein an, daß die ältesten Werkzeuge nicht aus Stein, sondern eher aus tierischem und pflanzlichen Material erarbeitet worden sein könnten und daß daher die Steinzeit, auch die Altsteinzeit, ein verhältnismäßig spätes Entwicklungsstadium des "Urmenschen" gewesen wäre; denn zuerst und unmittelbar lebe der Mensch in dem feinen Schleier von Wasser, Erde, Pflanze und Tier, der, umspült vom Luftmeer und dünn, den harten Steinkörper der Erde umgibt. Diesem harten Körper stehe jener Schleier, zu dem der Mensch selbst gehört, als Einheit gegenüber, ausgesetzt dem Einfluß der Gestirne und von ihren Kräften bewegt. Wir Menschen sind, als Körper betrachtet, Ausdrucksform der Umbildungen dieses Schleiers; die Kultur aber ist der Ausdruck des in diesem Schleier bei seiner Beziehung zu einem "Jenseits" symbolisch sich auswirkenden Lebens. Demnach, so meint wohl Frobenius, hat die älteste, die erste Kulturregung zu ihrer Gestaltung unwillkürlich nach den Elementen und Teilen des Schleiers selbst gegriffen, nicht nach dem Material der nackten, vom Schleier entblößten Steinkugel. So erwartet er noch die Aufdeckung einer viel älteren vorsteinzeitlichen Kultur, und die müßte vor der Quartärepoche, also in der Tertiärzeit liegen.
Bisher haben es nur
wenige Naturforscher gewagt, das Alter des
kulturfähigen Menschenwesens in tertiäre, ja in
vortertiäre Zeiten zurückzudatieren; so vor allem Hermann
Klaatsch. Seine eingehende und frei ausblickende anatomische
Analyse des menschlichen Körpers hat ihn schließlich ein
weit höheres erdgeschichtliches Alter des Menschen fordern lassen,
als man es bis dahin, befangen im Schematismus der älteren
Abstammungslehre, den Mut hatte anzunehmen. Noch in einer letzten
Arbeit (3) zu dieser Frage
sagt der verstorbene Anatom Schwalbe, Klaatschs Widerpart, daß
nach den ihm bekannten Tatsachen der Mensch vom Charakter des
javanischen Pithecanthropus
und des Homo Heidelbergensis
(Fig. 6g) erst in der allerletzten Phase der Tertiärzeit
entstanden sein könne.
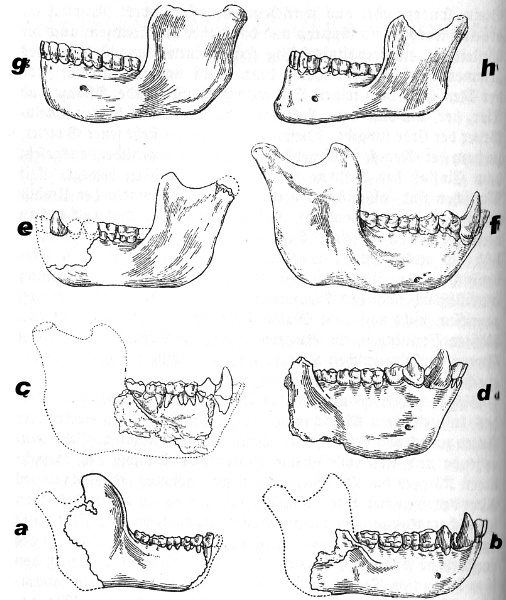
Unterkiefer von Affen und Menschen des känozoischen Zeitalters.
a) Propiopitheus. Älteste Affenform aus der Frühtertiärzeit (Oligozän). Ägypten. Eckzahnspitze abgebrochen. Angebliche Ahnenform aller Affen- und Menschenarten. 2/3 nat. Gr.
b) Piopithecus. Gibbon-ähnliche Form. Jüngere Tertiärzeit (Miozän). Deutschland. 2/3 nat. Gr.
c) Sivapithecus. Gorilla-ähnliche Form. Ende der Tertiärzeit (Pliozän). Indien. Stark verkl.
d) Dryopithecus. Schimpansen-ähnliche Form. Ende der Tertiärzeit (Pliozän). Süddeutschland. 1/3 nat. Gr.
e) Eoanthropus. Vermutlicher Menschenkiefer mit Affenmerkmalen. Jüngste Tertiärzeit (Pliozän). England. Der Eckzahn ist isoliert zugleich gefunden und hinkombiniert. 1/3 nat. Gr.
f) Orang Utan. Lebend. Jetztzeit. Indischer Archipel. 1/2 nat. Gr.
g) Homo Heidelbergensis. Diluvialzeit Eiszeitmensch. Mauer bei Heidelberg. Kieferform affenartig, Zähne menschenartig. 1/3 nat. Gr.
h) Homo sapiens. Lebend. Man beachte das vortretende Kinn, worin der Diluvialmensch affenähnlicher ist. 1/3 nat. Gr.
(Zusammengestellt aus W. K. Gregory, Journ. Dental. Research. 1920/21.)
Forscht man nun
aber in Schwalbes eigenen Darstellungen nach, welche
Glieder vergleichend anatomisch den Menschen mit niederen Formen, also
insbesondere mit den Affen nachweislich verbinden, so bekommt man
zusammenfassend die Antwort, daß zwar an der allgemeinen
tierischen Abstammung des Menschen nicht zu zweifeln, die spezielle
Abstammung von bestimmten Primatenaffen aber noch nicht gesichert
sei. Am wahrscheinlichsten sei die Abstammung des Menschen von
Formen mit einer Mischung der Charaktere niederer Ostaffen und
Menschenaffen. Diese Formen sind nun hypothetisch. Von
ihnen gehe der Weg zu einer ausgestorbenen - auch wieder hypothetischen
- Menschenaffenform, aus der die zum Schimpansen aufsteigende "Linie",
also noch nicht dieser selbst, sondern seine hypothetischen
Vorläufer sich entwickelten. "Aus dieser Linie hat sich dann
vermutlich der Stamm der zum Menschen führenden Hominiden
abgezweigt."
Wie kann man
angesichts eines so lustigen, doch nur aus imaginären
Formen zusammengesetzten Stammbaumes, der also das ganze Ergebnis
vieljähriger, streng an die vergleichende Anatomie gebundener
Forschung ist, noch behaupten, daß wir über die Entstehung
des Menschen aus Primaten im Klaren seien und daß andere, doch
auch mit sehr gewichtigen Beweisstücken arbeitende Theorien
über ein wesentlich höheres Alter des Menschenstammes und
über seine ursprüngliche Selbständigkeit gegenüber
allen bisher bekannt gewordenen vorweltlichen und jetztweltlichen
Säugetierformen keine Berechtigung haben sollten?
Noch können
wir nach Klaatsch (4)
die Zeit der Menschenwerdung in keiner Weise festlegen, wohl aber die
Tatsache festhalten, daß dieser Vorgang keinesfalls jüngeren
geologischen Datums ist. Früher wollte man die ganze
Entwicklung des Menschen bzw. der Menschenaffen aus niederen
Säugetieren in die Diluvialzeit oder in die unmittelbar ihr
vorausgehende letzte Phase der Tertiärzeit verlegen. Schon
der Nachweis, daß die hypostasierte Entwicklungsbahn Affe-Mensch
wohl teilweise umgekehrt verlief (5),
ferner die Erkenntnis, daß schon der Steinzeitmensch in mehrere
deutlich getrennte ältere Rassen zerfiel; drittens die
entwicklungsgeschichtliche Tatsache, daß alle Typen weit in die
Erdgeschichte zurückreichen; endlich die Erkenntnis, daß
Stammbäume sich nicht in dem äußerlichen Sinn entwerfen
lassen, wie es das Zeitalter der Deszendenztheorie in der zweiten
Hälfte des 19. Jahrhunderts glaubte - das alles hat den Boden
bereitet für die Erwartung einer sehr alten und noch durchaus
unaufgehellten Herkunft des Menschengeschlechtes. Mit Klaatsch
sehen wir heute dieses Problem so an: Der Tertiärmensch bedarf
nicht mehr des Beweises, einerlei, ob wir Artefakte von ihm haben und
Skelettfunde oder nicht; es kann sich nur noch um die Frage handeln, in
welcher Phase der Tertiärzeit die Trennung der Menschenaffen vom
Menschen einsetzte und wie alt der gemeinsame Stamm beider ist, der
nach Klaatschs wohlbegründeter Lehre unbedingt in das mesozoische
Erdzeitalter zurückreicht.
Die uns aus der Tertiärzeit bekannten Affengattungen (Fig. 6) sind so spezialisiert, daß sie, auch nach den bisherigen Forschungsmaximen der Abstammungslehre, überhaupt nicht als Ahnen eines Diluvialmenschen in Betracht kommen. Die als Ahnen sämtlicher Simiiden und Hominiden befürworteten (6) unzureichenden Schädelreste sind teils wegen ihrer Unvollständigkeit und dem Fehlen aller übrigen Skeletteile nicht auswertbar, teils sind es konvergente Formbildungen eigener Typenkreise. Der an Größe zurücktretende Eckzahn unterscheidet den Menschen ohnedies von allen echten Affen, die wir kennen, oder läßt höchstens deren Ableitung aus ihm selber zu. Doch ist die Eckzahnfrage in ein neues Stadium getreten, seit es durch Adolff wahrscheinlich gemacht wurde, daß Menschen- und Affeneckzahn gar keine homologen Bildungen sein müssen, daß sich vielleicht Verschiebungen im Gebiß schon beim frühesten Auseinandertreten beider Stammformen eingestellt haben, und daß der menschliche Eckzahn gegenüber dem der anthropoiden Affen so primitiv geblieben ist, daß deshalb eine Ableitung des Menschen von solchen Affen nicht angenommen werden kann. Selbst wenn frühere Vorfahren des Menschen einmal größere Eckzähne besessen haben sollten, würden das ganz ursprüngliche, aber gewiß keine Affeneckzähne gewesen sein (6a).
Die uns aus der Tertiärzeit bekannten Affengattungen (Fig. 6) sind so spezialisiert, daß sie, auch nach den bisherigen Forschungsmaximen der Abstammungslehre, überhaupt nicht als Ahnen eines Diluvialmenschen in Betracht kommen. Die als Ahnen sämtlicher Simiiden und Hominiden befürworteten (6) unzureichenden Schädelreste sind teils wegen ihrer Unvollständigkeit und dem Fehlen aller übrigen Skeletteile nicht auswertbar, teils sind es konvergente Formbildungen eigener Typenkreise. Der an Größe zurücktretende Eckzahn unterscheidet den Menschen ohnedies von allen echten Affen, die wir kennen, oder läßt höchstens deren Ableitung aus ihm selber zu. Doch ist die Eckzahnfrage in ein neues Stadium getreten, seit es durch Adolff wahrscheinlich gemacht wurde, daß Menschen- und Affeneckzahn gar keine homologen Bildungen sein müssen, daß sich vielleicht Verschiebungen im Gebiß schon beim frühesten Auseinandertreten beider Stammformen eingestellt haben, und daß der menschliche Eckzahn gegenüber dem der anthropoiden Affen so primitiv geblieben ist, daß deshalb eine Ableitung des Menschen von solchen Affen nicht angenommen werden kann. Selbst wenn frühere Vorfahren des Menschen einmal größere Eckzähne besessen haben sollten, würden das ganz ursprüngliche, aber gewiß keine Affeneckzähne gewesen sein (6a).
Vollends die
Halbaffen oder Lemuriden stehen durch ihre ganz
andersartigen und primitiven Merkmale einer formalen hypothetischen
Urform der Säugetiere näher als alle anderen
tertiärzeitlichen Säuger. Auch der
quartärzeitliche Mensch hat mehrere solcher ganz primitiven
Eigentümlichkeiten, welche ihn gleichfalls an hypothetische und
sehr weit in die Erdgeschichte zurückzudatierende
Vierfüßler anschließen. So ein lückenloses
vollständiges Gebiß, das nicht, wie das aller
tertiärzeitlichen Säugetiere, auf eine Lückenbildung
zwischen Vorder- und Backenzähnen hinläuft, sondern sich
allenfalls nur bei uns Spätmenschen in einem Ausfall des letzten
Backenzahns etwas verringert. Es gleicht in seiner
Vollständigkeit den ältesten tertiärzeitlichen
Säugetierformen; aber an solche den Menschen anzuschließen,
verbieten die Grundsätze der bisherigen Deszendenzlehre selbst,
wonach voneinander durch einseitige Differenzierung schon
unterschiedene Lebewesen nicht unmittelbar stammesgeschichtlich
zusammenhängen können. Auch in der fünffingerigen
Extremität liegt ein sehr altes, nicht ein erdgeschichtlich
jungzeitliches Merkmal, das keineswegs die Ableitung des Menschentypus
aus irgend einem tertiärzeitlichen Säugetier
zuläßt. Zwar gibt es alttertiäre Säugetiere
mit der primitiven fünffingerigen Extremität; aber sie, wie
auch spätere Gattungen mit demselben Merkmal, sind so einseitig
entwickelt (7),
daß man die Herkunft der fünffingerigen
Primatenextremität, insbesondere der vorderen, in viel ältere
Zeiten zurückdatieren kann. Sie ist in der beim Menschen
vorhandenen Einfachheit schon paläozoischen Alters. Zwar ist
es ein Irrtum, die vollständig fünffingerige Extremität
für den Urzustand des Landtieres anzusehen, wie man es bisher
tat. Neueste Funde und Untersuchungen fossiler Uramphibien aus
der Permzeit haben ergeben, daß schon jene frühen
amphibischen Landtiere eine reduzierte, nämlich eine vierzehige
Vorderextremität hatten, während ihre Hinterextremität
noch fünfzehig war. Sind sie in dieser Hinsicht schon
einseitig spezialisierte Formen gewesen, so stehen sie auch dem
primitiven hypothetischen Landtier insofern schon ferner, als dieses
wahrscheinlich zuerst siebenzehig war. Nicht nur die
Embryonaluntersuchung gewisser lebender Amphibien ergibt dies, sondern
es erhellt auch aus Fossilfunden permzeitlicher alter Amphibienformen,
deren noch fünfzehige Hinterextremität die Reste eines
überzähligen, jedoch nicht pathologischen oder atavistischen,
sondern noch als Normalrest vorhandenen Kleinfingers enthält (8). Es war demnach
die fünffingerige Landextremität schon gegen Ende der
paläozoischen Zeit selbst ein Rückbildungsstadium aus einer
noch älteren Form. Wir können also festhalten,
daß mindestens die fünffingerige Extremität ein
Grundmerkmal aller zum erstenmal völlig dem Leben auf dem Festland
zugewandten, wenn auch habituell noch amphibisch aussehenden Wesen war;
daß mithin die fünffingerige Landextremität mindestens
ein Erbe aus der Steinkohlenzeit ist. Und so kann der Mensch mit
allen gleichfalls fünffingerigen Säugetieren bis in jene alte
Epoche zurückreichen, weil alle bis dorthin fossil bekannt
gewordenen Vierfüßler in ihren Extremitäten oder im
übrigen Körper schon viel zu einseitig entwickelt sind, als
daß die Ableitung des menschlichen Körpers aus ihnen
gelänge.
Die Frage, ob und
wie die Hand des Menschen aus einem allenfalls
anzunehmenden Urextremitätenstadium hervorgegangen sein
könnte, ist weder vergleichend anatomisch noch durch Fossilfunde
zu beantworten. Auch Klaatsch hat sich mit dieser
Grenzfeststellung unseres vergleichend anatomischen Wissens
begnügt und sagt nur, daß für die Betrachtung der
Herkunft des Menschen die Feststellung vorerst ausreicht, daß er
die Hand mit der fünffingerigen Form der alten Landwirbeltiere,
also nicht erst mit jener der tertiärzeitlichen Säugetiere
teilt. Auch dürfte nun klar sein, sagt Klaatsch, daß
die früher vielfach übliche Einschätzung des Menschen
als des letzten Endgliedes einer angeblich mühsamen Entwicklung
nicht zutrifft. Er ist nicht als letztes Ergebnis einer sehr
komplizierten Umgestaltung aufzufassen. Es fehlte früher
nicht an solchen Vermutungen, als man glaubte, daß der Mensch von
vierfüßigen Tieren abstammen könne. Aber ein
Vierfüßlertum besagt, daß die Gliedmaßen des
betreffenden Wesens zu Füßen gestaltet sind, d. h.
außer Stützen und Laufen andere Leistungen nicht verrichten
können. Der Mensch aber habe das alte vielseitige und daher
unendlich wertvolle Werkzeug beibehalten. "Daß der Mensch indifferent blieb,
sich seine Vielseitigkeit bewahrte - darin liegt eben ein großer
Teil des Geheimnisses seines außerordentlichen Erfolges.
Das ist eine ganz andere Auslegung des Entwicklungsganges als die im
alten Darwinschen Sinne. Nicht ein Triumph des Kampfes ums Dasein
ist der Mensch; nein, im Gegenteil: sein Sieg beruht darin, daß
er von den Opfern der natürlichen Zuchtwahl verschont blieb,
daß er seine Hand behielt. Wenn man nach Wundern suchen
will, so braucht man nicht ins Reich des Übernatürlichen zu
flüchten. Die Natur selbst, unser eigenes Wesen, bietet der
Wunder genug. Nach den Regeln der Wahrscheinlichkeitsordnung ist
jedenfalls die Ausnahmestellung des Menschen gegenüber dem ganzen
anderen Tierreich eine sehr sonderbare. Nicht der Besitz der Hand
als solcher ist es - er war ja allen Tieren einstmals eigen - sondern
der Umstand, daß dieses Gebilde in seiner Ursprünglichkeit
beibehalten wurde und daß es sich in den Dienst einer gewaltigen
Gehirnentfaltung stellen konnte - das ist das Merkwürdige."
Der Mensch kann
auch deshalb mindestens mesozoischen, ja
paläozoischen Alters sein, weil er ein Sohlengänger
ist. Die ältesten, hypothetischen und die aus ihnen formal
allenfalls ableitbaren tertiärzeitlichen Säugetiertypen sind
vollkommene Sohlengänger oder weisen wenigstens als
Abkömmlinge auf Formen mit Sohlengängerfuß eindeutig
hin. Der Mensch selbst kann wiederum nicht von solchen Gattungen
abstammen, sondern hatte bestenfalls in sehr früher, dem
paläozoischen Ursprungspunkt der Säugetiere recht nahen Zeit
einen dort schon zu suchenden Ahnen mit ihnen. Somit hat er
mindestens seit dem Altmesozoikum auf eigener Entwicklungsbahn den
Sohlengänger in sich bewahrt, was nicht hindert, daß seine
große Zehe am Fuß zuerst opponierbar war, der Fuß
also auch Greifhandcharakter besaß. Er war somit auch in
mesozoischer Zeit schon ein von den späteren
tertiärzeitlichen Stämmen getrennter eigener
Säugetierstamm, mithin als Menschentypus vorhanden, wenn auch in
anderer Form als zur Steinzeit.
Klaatsch hat seinerzeit auch gewisse Landtierfährten im mitteldeutschen Sandstein aus frühmesozoischer Zeit zu einem Analogiebeweis für das hohe Alter des Menschentypus herangezogen und ich folgte ihm ursprünglich darin. Sie gehören sehr alten Landtieren an, die man nur aus diesen Spuren, nicht nach Skelettresten kennt (Fig. 7).
Klaatsch hat seinerzeit auch gewisse Landtierfährten im mitteldeutschen Sandstein aus frühmesozoischer Zeit zu einem Analogiebeweis für das hohe Alter des Menschentypus herangezogen und ich folgte ihm ursprünglich darin. Sie gehören sehr alten Landtieren an, die man nur aus diesen Spuren, nicht nach Skelettresten kennt (Fig. 7).

Fig. 7 (Bildquelle/-text: "Urwelt, Sage und
Menschheit"
von Edgar Dacqué, 8. Aufl. 1938, R. Oldenbourg)
Fährten eines Reptils (Chirotherium) aus dem altmesozoischen Thüringer Buntsandstein. Scheinbar mit Daumen. Die Hinterextremität größer,
die Vorderextremität kleiner. 1/6 nat. Gr. (Nach R. Owen aus Zittel-Broili, Grundzüge der Paläontologie II. 1923.)
Fährten eines Reptils (Chirotherium) aus dem altmesozoischen Thüringer Buntsandstein. Scheinbar mit Daumen. Die Hinterextremität größer,
die Vorderextremität kleiner. 1/6 nat. Gr. (Nach R. Owen aus Zittel-Broili, Grundzüge der Paläontologie II. 1923.)
Sie fallen auf
durch ihre große Ähnlichkeit mit
Menschenhänden, wenn sie auch solchen gewiß nicht
zugehörten. Die älteren, aus der Permzeit, zeichnen
sich dadurch aus, daß die gegenseitigen
Größenverhältnisse der vorderen und hinteren
Extremitäten sich noch mehr angleichen als die der etwas
jüngeren aus der Triaszeit, wo die hinteren Extremitäten die
vorderen an Größe wesentlich übertreffen. Aus der
Form der Abdrücke kann man den Schluß ziehen, daß das
zugehörige "Handtier" (Chirotherium), wenn auch nicht ganz
aufrecht, so doch zuerst weniger, später entschiedener auf der
Hinterextremität lief. Besonders die älteren
Fährten seien deshalb bedeutungsvoll, weil gerade sie große
Ähnlichkeit mit Kinderhänden haben. Man kann sie, sagt
Klaatsch, in der Tat nicht besser beschreiben als durch einen Vergleich
mit solchen oder auch mit Embryonalzuständen der Menschenhand
(Fig. 8), worin die Plumpheit und Derbheit der Kinderhand noch
auffälliger erscheint.

Fig. 8 (Bildquelle/-text: "Urwelt, Sage und
Menschheit"
von Edgar Dacqué, 8. Aufl. 1938, R. Oldenbourg)
Embryonalhand des Menschen. (Aus Klaatsch, Abstammungslehre. 1911.)
Embryonalhand des Menschen. (Aus Klaatsch, Abstammungslehre. 1911.)
Ein solches
embryonales Merkmal der Menschenhand, wie es auch den
Thüringer Fährten eignet, ist die wenig ausgedehnte
Fläche zwischen dem Daumenballen und den übrigen Fingern,
oder die außerordentlich breite Mittelhand oder die kurzen
gedrungenen Finger. Der Daumen an der Thüringer Spur ist mit
einem dicken Ballen versehen und bekundet damit seine
Opponierbarkeit. Die Unterfläche der Finger zeigt oft die
Gelenkvertiefungen, den Fingergliedern entsprechend. Der vierte
Finger war meistens am längsten.
Diese Auffassung ist nicht mehr haltbar, seitdem man durch neuere grundlegende Untersuchungen Soergels weiß (8a), was man auch früher teilweise schon gemutmaßt hatte, daß der "Daumen" an diesen Extremitäten gar kein Daumen ist, sondern an der entgegengesetzten Seite der "Hand" liegt, mithin eine sekundäre Erwerbung, eine "falsche" Zehe darstellt und so als Anpassungsbildung an das Aufrechtgehen zur Unterstützung der Sohle oder als Verbreiterung der Fußbasis zur besseren Verhütung des Einsinkens vielleicht gedeutet werden kann. Dagegen sind diese Formen ein erneuter Beweis für das Herrschen einer Formbildung, die auch in anderen mesozoischen Typen mit der Tendenz zum Halbaufrechtgehen zusammentrifft und dabei eine gewisse Spreizbarkeit eines Zehengliedes hervorbringt.
Überhaupt finden wir bei den Landtieren des mesozoischen Zeitalters, wie früher schon erwähnt, die Anlage zum aufrechten Gang auf den Hinterbeinen und dies oft in einer so volkommenen Weise, daß es geradezu als Zeitmerkmal angesprochen werden darf. Damit verknüpft ist merkwürdigerweise gelegentlich der opponierbare Daumen, der auch der Verteidigung gedient haben soll. Zeigten uns dies schon in der schönsten Art die Sandsteinfährten, so haben wir auch ganze Skelette verschiedenster Reptilgruppen der mesozoischen Epoche, besonders der Kreidezeit und der oberen Jurazeit, welche jenes Kennzeichen ganz vollkommen entwickelt haben. So etwa die aufrechten Iguanodonten der belgischen Unterkreideformation. Es waren das bis 5 m hohe landbewohnende Pflanzenfresser, habituell sehr ähnlich anderen, z. B. nordamerikanischen und afrikanischen Landsauriern mit Raubtier- wie mit Pflanzenfressercharakter, sehr großer und kleiner Dimension. Sie zeigen, wie schon erwähnt, den aufrechten Gang, haben aber nur eine dreizehige oder höchstens vierzehige Extremität, an der das fünfte Glied allenfalls noch als Rudiment hängt. Dagegen hatten gerade nicht die jüngeren, sondern die erdgeschichtlich ältesten Landtiere des paläozoischen Zeitalters entschiedene Anlage zu einer opponierbaren ersten Zehe, unserem menschlichen Daumen entsprechend. Und dies ist umsomehr ein Beweis für die Altertümlichkeit des Menschentypus, als die erdgeschichtlich jüngeren, alttertiärzeitlichen Säugetiere zwar vielfach noch vollkommen fünffingerige Extremitäten besaßen, aber dabei keinen opponierbaren Daumen hatten und außerdem auch zu Gruppen gehörten, deren spezifische Ausbildung sie gewiß für keinen Forscher zu einem stammesgeschichtlichen Anknüpfungspunkt gerade des Menschen oder menschenähnlicher Primaten werden läßt. So geht also die Menschenhand in ihrer Grundlage mindestens auf älteste Landtierformen zurück, wenn wir auch den Menschen nicht aus jenen Altformen stammesgeschichtlich herleiten möchten. Denn der Mensch ist vergleichend anatomisch zunächst ein Säugetier, kein Reptil, und ist grundsätzlich typenhaft ein Vierhänder, kann also auch nicht von Vierfüßlern "abstammen". Sie zeigen uns aber immerhin, in welcher Epoche uralter Formbildung seine Extremitäten ideell wurzeln.
Ebenso weisen die im vorigen Abschnitt beschriebenen Landreptilien des Erdmittelalters mit ihrem aufrechten Gang und ihrer einfach gestreckten Hinterextremität, deren Länge nicht wie bei spätzeitlichen Säugetieren erst sekundär durch Aufrichtung des Fußes (9) bewirkt ist, auf eine mesozoische Zeitsignatur hin, und wir finden den Menschen auch mit diesem Merkmal ausgestattet. Auch dies deutet vergleichsweise auf sein hohes erdgeschichtliches Alter hin.
Diese Auffassung ist nicht mehr haltbar, seitdem man durch neuere grundlegende Untersuchungen Soergels weiß (8a), was man auch früher teilweise schon gemutmaßt hatte, daß der "Daumen" an diesen Extremitäten gar kein Daumen ist, sondern an der entgegengesetzten Seite der "Hand" liegt, mithin eine sekundäre Erwerbung, eine "falsche" Zehe darstellt und so als Anpassungsbildung an das Aufrechtgehen zur Unterstützung der Sohle oder als Verbreiterung der Fußbasis zur besseren Verhütung des Einsinkens vielleicht gedeutet werden kann. Dagegen sind diese Formen ein erneuter Beweis für das Herrschen einer Formbildung, die auch in anderen mesozoischen Typen mit der Tendenz zum Halbaufrechtgehen zusammentrifft und dabei eine gewisse Spreizbarkeit eines Zehengliedes hervorbringt.
Überhaupt finden wir bei den Landtieren des mesozoischen Zeitalters, wie früher schon erwähnt, die Anlage zum aufrechten Gang auf den Hinterbeinen und dies oft in einer so volkommenen Weise, daß es geradezu als Zeitmerkmal angesprochen werden darf. Damit verknüpft ist merkwürdigerweise gelegentlich der opponierbare Daumen, der auch der Verteidigung gedient haben soll. Zeigten uns dies schon in der schönsten Art die Sandsteinfährten, so haben wir auch ganze Skelette verschiedenster Reptilgruppen der mesozoischen Epoche, besonders der Kreidezeit und der oberen Jurazeit, welche jenes Kennzeichen ganz vollkommen entwickelt haben. So etwa die aufrechten Iguanodonten der belgischen Unterkreideformation. Es waren das bis 5 m hohe landbewohnende Pflanzenfresser, habituell sehr ähnlich anderen, z. B. nordamerikanischen und afrikanischen Landsauriern mit Raubtier- wie mit Pflanzenfressercharakter, sehr großer und kleiner Dimension. Sie zeigen, wie schon erwähnt, den aufrechten Gang, haben aber nur eine dreizehige oder höchstens vierzehige Extremität, an der das fünfte Glied allenfalls noch als Rudiment hängt. Dagegen hatten gerade nicht die jüngeren, sondern die erdgeschichtlich ältesten Landtiere des paläozoischen Zeitalters entschiedene Anlage zu einer opponierbaren ersten Zehe, unserem menschlichen Daumen entsprechend. Und dies ist umsomehr ein Beweis für die Altertümlichkeit des Menschentypus, als die erdgeschichtlich jüngeren, alttertiärzeitlichen Säugetiere zwar vielfach noch vollkommen fünffingerige Extremitäten besaßen, aber dabei keinen opponierbaren Daumen hatten und außerdem auch zu Gruppen gehörten, deren spezifische Ausbildung sie gewiß für keinen Forscher zu einem stammesgeschichtlichen Anknüpfungspunkt gerade des Menschen oder menschenähnlicher Primaten werden läßt. So geht also die Menschenhand in ihrer Grundlage mindestens auf älteste Landtierformen zurück, wenn wir auch den Menschen nicht aus jenen Altformen stammesgeschichtlich herleiten möchten. Denn der Mensch ist vergleichend anatomisch zunächst ein Säugetier, kein Reptil, und ist grundsätzlich typenhaft ein Vierhänder, kann also auch nicht von Vierfüßlern "abstammen". Sie zeigen uns aber immerhin, in welcher Epoche uralter Formbildung seine Extremitäten ideell wurzeln.
Ebenso weisen die im vorigen Abschnitt beschriebenen Landreptilien des Erdmittelalters mit ihrem aufrechten Gang und ihrer einfach gestreckten Hinterextremität, deren Länge nicht wie bei spätzeitlichen Säugetieren erst sekundär durch Aufrichtung des Fußes (9) bewirkt ist, auf eine mesozoische Zeitsignatur hin, und wir finden den Menschen auch mit diesem Merkmal ausgestattet. Auch dies deutet vergleichsweise auf sein hohes erdgeschichtliches Alter hin.
Im
Spätpaläozoikum, in der Permzeit sich andeutend, besonders
aber in der Triaszeit klar hervortretend, stoßen wir auf eine
Erscheinung, die uns offenbart, wann der Säugetiertypus als
Zeitsignatur erwachte und sich vollendete. Da gibt es auf einem
großen, später untergegangenen Südkontinent, sowie
teilweise in Nordamerika, etwas später auch in Rußland
erscheinend, eine Reptilgruppe, die Theromorphen, denen
auffallenderweise in vielen ihrer Geschlechter Organe und ein
allgemeiner Habitus von Säugetiercharakter zuteil wird.
Unter ihnen treten sogar Formen auf mit säugetierartigem Raubtier-
und Pflanzenfressergebiß, wie sie echte tertiär- und
jetztzeitliche Säugetiere haben (Fig. 9); aber ihrem inneren Bau
und Wesen nach sind diese scheinbaren Säugetiermerkmale eben doch
reptilhaft gewesen.
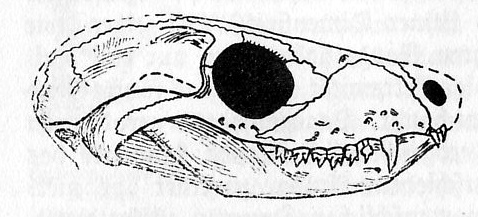
Fig.
9 (Bildquelle/-text:
"Urwelt, Sage und Menschheit" von Edgar Dacqué, 8. Aufl. 1938,
R.
Oldenbourg)
Schädel eines Reptils der Triaszeit, mit Säugetiermerkmalen und Säugetiergebiß. Südafrika.
(Aus W.K. Gregory. Journ. Dental. Research. 1920.) Nat. Gr.
Schädel eines Reptils der Triaszeit, mit Säugetiermerkmalen und Säugetiergebiß. Südafrika.
(Aus W.K. Gregory. Journ. Dental. Research. 1920.) Nat. Gr.
Auch das Skelett
bleibt in entscheidenden Zügen das unverkennbare
Reptilskelett, wenn die äußere Gestaltung und Formung oft
aufs stärkste an das Säugetier erinnert. Unter dem
Einfluß der formalistischen Deszendenzlehre hat man immer und
immer wieder versucht, jene Gestalten als die leibhaftigen Ahnen der
späteren Säugetiere anzusprechen; doch ist das jedesmal
mißglückt. Wir haben eben hier wieder die einander
formal überschneidenden Formenkreise nach dem gegebenen Schema
(Fig. 2B). In der Permzeit ist der Augenblick eingetreten, wo das
Säugetierhafte gewissermaßen in der Luft liegt, wo es
beginnt, Zeitsignatur zu werden, so daß eben auch ein
großer Teil der Reptilien jenes Zeitalters den
Säugetierhabitus annimmt, ohne je Säugetier gewesen zu sein
oder stammesgeschichtlich überhaupt in jene Bahn
einzulenken. Wenn man unter dieser Zeitgesellschaft, wie zu
erwarten, einmal echte einwandfreie Säugetierahnen und damit also
Säugetiere selbst finden sollte, so werden sie einigen
theromorphen Reptilien ebenso ähnlich sein, wie etwa die
tertiärzeitlichen Affen dem Menschen: man hat es dann auch wieder
mit formalen Überschneidungen der festen Typenkreise zu tun, deren
Deckungsflächen eine natürliche Familie vortäuschen und
Arten liefern, die wie Stammarten oder Urformen der übrigen, sich
formal anschließenden Gattungen, sowohl der Reptilien wie der
Säugetiere, aussehen werden und es doch nicht sind.
Hiermit ist der
Zeitpunkt ermittelt, in dem mindestens das früheste
Säugetierwerden vor sich ging. Dort muß der
Menschenstamm mit seiner Säugetiernatur und seiner
fünffingerigen, sohlengängerigen Extremität mindestens
wurzeln - als säugetierhafter Menschenstamm nämlich,
unterscheidbar von anderen, gleichzeitig mit ihm erscheinenden
Typen. Niemals aber
können Reptilien Säugetiere geworden sein. Ihr
Skelettbau und andere Merkmale, wie das Fehlen der Hautdrüsen,
läßt das nicht möglich erscheinen. Wir
müssen, wenn wir überhaupt Stammbaumversuche machen wollen,
dem permischen hypothetischen Säugetiertypus amphibische Merkmale
beilegen, nicht weil er etwa aus echten Altamphibien hervorgegangen
wäre, sondern weil damals die amphibische Gestalt und Lebensweise
eine Zeitsignatur war, dementsprechend das Ursäugetier jenes
Gewand trug. Da nun die ältesten Säugetiere, die wir
nach Zahnfunden aus der Triaszeit kennen, schon einseitig als
Beuteltiere spezialisiert sind, so daß sie sich schon
frühzeitig vom übrigen Säugetier- und Menschenstamm
getrennt haben mußten; da ferner das Reptil nicht in die
Stammbahn gehört haben kann, so muß der säugetierhafte
Menschenstamm in der Permzeit amphibienhafte äußere Merkmale
gehabt haben, aber schon damals als solcher selbständig
aufgetreten sein.
Ganz folgegerecht ist also der Schluß, zu dem schon Klaatsch in Würdigung der menschlichen Anatomie gelangte, ohne sich indessen der ganzen ungeheuren Tragweite seiner Entdeckung bewußt geworden zu sein, wenn er sagt: "Die ältesten Stadien der menschlichen Vorgeschichte werden daher mit denjenigen der Urgeschichte der Landwirbeltiere identisch sein." Diese Urgeschichte aber liegt schon im paläozoischen Zeitalter.
Ganz folgegerecht ist also der Schluß, zu dem schon Klaatsch in Würdigung der menschlichen Anatomie gelangte, ohne sich indessen der ganzen ungeheuren Tragweite seiner Entdeckung bewußt geworden zu sein, wenn er sagt: "Die ältesten Stadien der menschlichen Vorgeschichte werden daher mit denjenigen der Urgeschichte der Landwirbeltiere identisch sein." Diese Urgeschichte aber liegt schon im paläozoischen Zeitalter.
Der einzige
Paläontologe, welcher die nächstliegende
Folgerung aus den neueren stammesgeschichtlichen und vergleichend
anatomischen Erkenntnissen auch für den Menschenstamm gezogen hat,
wenngleich in noch wenig gegenständlicher Umreißung, ist
Steinmann gewesen (10).
Bei der Langsamkeit der Entwicklung aller Tierstämme, sagt er,
dürfe man erwarten, daß sich auch die Menschwerdung im Stamm
der Anthropotherien außerordentlich langsam vollzogen habe.
Weil Skelettreste und Werkzeuge des Menschen erst aus der Diluvialzeit
bekannt geworden seien, halte man den Menschen als solchen erst
für ein Erzeugnis dieses letzten erdgeschichtlichen
Abschnittes. Wahrscheinlicher sei es, daß der Mensch,
ebenso wie die ihn zur Diluvialzeit begleitenden Tiere: Pferd, Elefant,
Nashorn, Nilpferd, Hirsch usw. schon zur Tertiärzeit existierte,
wenn auch in einem etwas anderen, stammesgeschichtlich
altertümlichen Gewand. Wie man am Anfang der Stammbahnen nie
die von der älteren Deszendenzlehre erwarteten generalisierten
Grundformen gefunden habe, aus denen man nachfolgende Spezialtypen
ableiten könne, so kenne man aus dem ältesten Abschnitt der
Tertiärzeit nicht weniger als 25 Gattungen des Primatentypus, von
denen keine die Forderungen erfülle, die sie in anatomischer
Hinsicht zu einem stammesgeschichtlichen Bindeglied zwischen Mensch und
niederen Primaten machte. Offenbar haben also beide Typen schon
am Anfang der Tertiärzeit eine lange selbständige Geschichte
hinter sich, so daß man dem Schluß nicht mehr ausweichen
könne, daß die ganze Sippe in mehreren getrennten Linien in
das mesozoische Zeitalter zurückreiche.
Klaatsch und
Steinmann haben mit ihren Ideen und
Schlußfolgerungen etwas in der Hand gehabt, dessen ganze
Tragweite sie noch nicht erfaßten. Vielleicht hat es
Klaatsch, viel angefeindet und als Phantast verschrien, stärker
geahnt, doch drang es bei ihm jedenfalls nicht mehr zum vollen
Bewußtsein durch. Keiner von beiden hat es sich klar
gemacht und es als unerbittliche Konsequenz seiner Beweisführung
angesehen, daß er methodisch und im Resultat die ganze
menschliche und tierische Abstammungslehre, wie sie stereotyp seit
sechzig Jahren (Anm. der WFG: Heute, seit 140 Jahren)
ohne irgend einen wesentlich neuen, vertiefenden Gesichtspunkt
vertreten ward, völlig umgestürzt und die Typentheorie und
somit auch die Erkenntnis der Selbständigkeit des Menschenstammes
bis in älteste Zeiten des Landtierwerdens an deren Stelle gesetzt
hat.
Die Forschung ist
dabei nicht stehengeblieben. Seit der ersten
Herausgabe dieser Blätter haben sich durch Untersuchung innerer
und äußerer Organe noch andere bedeutsame Hinweise auf ein
sehr hohes Alter des Menschengeschlechtes und auf gewisse
Vorfahrenstadien ergeben (10a).
Die Kinnbildung bzw. Kieferbildung stellt einen Zustand dar, der
unmittelbar auf die uralte Einwärtsneigung der Zähne bei
Haifischen und gewissen Reptilien folgt, also an den frühesten
Beginn der Säugetierzeit und teilweise sogar noch an ein
Vorsäugetierstadium erinnert. Man könnte, sagt
Westenhöfer, den Versuch machen, einem Unbefangenen eine Reihe
verschiedener Unterkiefer zu übergeben, damit er den menschlichen
an die Stelle setze, wo er nach seiner ganzen Entwicklung
vergleichsweise hingehört; er würde ihn jedenfalls vor jedes Säugetier
stellen. Das aber heißt nichts anderes, als daß auch
in der Zahnstellung und Kieferbildung alle Säugetiere so
spezialisiert sind, daß der Mensch auch hierin ihr Ahnenstadium,
nicht ihr Nachkommenstadium ist.
Demselben Forscher verdanken wir die Erkenntnis, daß wir im Kindheitszustand, sowie ein nicht geringer Prozentsatz Erwachsener, mit dem trichterförmigen Wurmfortsatz des Blinddarmes, mit einer gelappten statt einer geschlossenen Niere und einer mehrfach gekerbten Milz auf einer stammesgeschichtlich tieferen Entwicklungsstufe stehengeblieben sind. Dieser dreifache "Progonismus" deutet merkwürdigerweise auf Wassertierstadien hin, indem jene Formbildungen der Niere und Milz nur von wasserangepaßten Säugern erreicht oder übertroffen werden. Und so ist die Möglichkeit nicht von der Hand zu weisen, daß in der spezifisch menschlichen Vorfahrenreihe eine aquatile Form vorhanden war. Westenhöfer meint, daß diese Mischung von progonischen und nichtprogonischen Menschen dadurch zustande gekommen sei, daß sie sich vielleicht aus zwei Stämmen mischten, von denen der eine schon etwas weiter fortgeschritten, der andere etwas zurückgeblieben war; dieser würde dem Menschenaffenvorfahren nähergestanden haben, jener schon jetztweltmenschlicher gewesen sein; beider Wurzeln lägen aber weit zurück in erdgeschichtlicher Vergangenheit.
Demselben Forscher verdanken wir die Erkenntnis, daß wir im Kindheitszustand, sowie ein nicht geringer Prozentsatz Erwachsener, mit dem trichterförmigen Wurmfortsatz des Blinddarmes, mit einer gelappten statt einer geschlossenen Niere und einer mehrfach gekerbten Milz auf einer stammesgeschichtlich tieferen Entwicklungsstufe stehengeblieben sind. Dieser dreifache "Progonismus" deutet merkwürdigerweise auf Wassertierstadien hin, indem jene Formbildungen der Niere und Milz nur von wasserangepaßten Säugern erreicht oder übertroffen werden. Und so ist die Möglichkeit nicht von der Hand zu weisen, daß in der spezifisch menschlichen Vorfahrenreihe eine aquatile Form vorhanden war. Westenhöfer meint, daß diese Mischung von progonischen und nichtprogonischen Menschen dadurch zustande gekommen sei, daß sie sich vielleicht aus zwei Stämmen mischten, von denen der eine schon etwas weiter fortgeschritten, der andere etwas zurückgeblieben war; dieser würde dem Menschenaffenvorfahren nähergestanden haben, jener schon jetztweltmenschlicher gewesen sein; beider Wurzeln lägen aber weit zurück in erdgeschichtlicher Vergangenheit.
Wie man sieht,
mehren sich die Hinweise auf die Altertümlichkeit
und Primitivität des Menschen. Nur in der Gehirnentwicklung
und damit im vollendeten Bau des Gesichtsschädels ist der Mensch
allen voraus, die Affen aber sind schon vor diesem vollendeten Stadium
seitlich abgewichen und haben sich degenerativ spezialisiert. Und
Westenhöfer sagt treffend, daß alle diese Tatsachen doch
wohl nichts anderes bedeuten können, als daß der
Säugetierstamm insgesamt den Weg zum Großhirn eingeschlagen
hatte, daß diese Entwicklung aber in umso stärkerem
Maße gehemmt wurde, je mehr sich das Gebiß neigte, d. h. je
stärker und mächtiger es zugleich wurde. In unsere
Ausdrucksweise übersetzt, bedeutet dies aber: die Säugetiere
sind auf dem Entwicklungsweg des Menschenstammes zurückgeblieben
und von der Abzweigungsstelle aus dann einseitig spezialisierte Ableger
des gemeinsamen und durchlaufenden Urstammes geworden, der zu allen
Zeiten eben der "Menschenstamm" selber war.
Doch wie dem auch sei, muß der uralte Stamm des Menschentypus - darüber kann es ja paläontologisch kaum eine grundsätzliche Meinungsverschiedenheit geben - in vergangenen Erdperioden ein anderes Aussehen gehabt haben, ob er nun aus irgend welchen Tierformen genetisch hervorbrach oder ob er von ältester Landtierzeit her selbständig seine Bahn verfolgte; er hat zweifellos seine Evolution gehabt und manche Mutationen hervorgebracht.
Aber welche? Wir erinnern uns jetzt noch einmal des oben dargelegten Gesetzes, wonach in bestimmten Zeitepochen bestimmte Gestaltungen und Organe bei den verschiedensten Typen zum Vorschein kommen. Hat aber, so sagten wir, eine Tiergruppe oder eine einzelne Gattung in späterer Zeit noch einen für eine frühere Zeit festgestellten zeitcharakteristischen Formzustand voll entwickelt oder rudimentär an sich, so erkennt man daran ihr geologisches Mindestalter, auch ohne daß man aus früheren Zeiten ihres Daseins fossile Dokumente von ihr hätte. Wenn also der Menschenstamm - das Wort meinetwegen in jedem beliebigen stammesgeschichtlichen Sinn gebraucht - noch in das paläozoische und mesozoische Zeitalter zurückgeht, so muß er eben solche Formeneigentümlichkeiten damals gehabt haben und jetzt noch voll entwickelt oder rudimentär an sich tragen; also z. B. den opponierbaren Daumen oder den aufgerichteten Gang auf primär, nicht sekundär verlängerten bzw. aufgerichteten Hinterbeinen. Und das hat er. Er hat aber auch in seiner Extremität den primitiven Formzustand des Landtieres, wenn auch etwas abgeändert, bewahrt, und der ist mindestens jungpaläozoischer Herkunft.
Doch wie dem auch sei, muß der uralte Stamm des Menschentypus - darüber kann es ja paläontologisch kaum eine grundsätzliche Meinungsverschiedenheit geben - in vergangenen Erdperioden ein anderes Aussehen gehabt haben, ob er nun aus irgend welchen Tierformen genetisch hervorbrach oder ob er von ältester Landtierzeit her selbständig seine Bahn verfolgte; er hat zweifellos seine Evolution gehabt und manche Mutationen hervorgebracht.
Aber welche? Wir erinnern uns jetzt noch einmal des oben dargelegten Gesetzes, wonach in bestimmten Zeitepochen bestimmte Gestaltungen und Organe bei den verschiedensten Typen zum Vorschein kommen. Hat aber, so sagten wir, eine Tiergruppe oder eine einzelne Gattung in späterer Zeit noch einen für eine frühere Zeit festgestellten zeitcharakteristischen Formzustand voll entwickelt oder rudimentär an sich, so erkennt man daran ihr geologisches Mindestalter, auch ohne daß man aus früheren Zeiten ihres Daseins fossile Dokumente von ihr hätte. Wenn also der Menschenstamm - das Wort meinetwegen in jedem beliebigen stammesgeschichtlichen Sinn gebraucht - noch in das paläozoische und mesozoische Zeitalter zurückgeht, so muß er eben solche Formeneigentümlichkeiten damals gehabt haben und jetzt noch voll entwickelt oder rudimentär an sich tragen; also z. B. den opponierbaren Daumen oder den aufgerichteten Gang auf primär, nicht sekundär verlängerten bzw. aufgerichteten Hinterbeinen. Und das hat er. Er hat aber auch in seiner Extremität den primitiven Formzustand des Landtieres, wenn auch etwas abgeändert, bewahrt, und der ist mindestens jungpaläozoischer Herkunft.
Mit dieser
Erkenntnis dürfen wir jetzt nach dem Gesetz der
Zeitcharaktere den Menschenstamm bis in das paläozoische
Erdzeitalter zurückführen und für seine Evolution
folgende Stadien annehmen: Zuerst muß er amphibische und
reptilhaft scheinende Merkmale besessen haben. Er hatte
vielleicht, wie die Amphibien, den schleppenden Gang und
schwimmhautartig verwachsene Finger und Zehen, auch wohl noch keinen
entschieden opponierbaren Daumen. Mit dem ältesten Amphibien
und Reptilien hatte er vielleicht einen teilweise hornig gepanzerten
Körper gemein, ein Merkmal, das überhaupt in der Erdzeit der
paläozoischen Epoche als Zeitcharakteristikum insofern gelten
kann, als auch die Amphibien mit ihrem an und für sich
schleimigen, drüsenbesetzten Hautmantel zu solcher Panzerbildung,
oft an Kehle und Brust, auch auf dem Rücken, bis zur Stärke
von Hautknochenplatten sich steigernd, übergehen. Mit beiden
Gruppen aber hatte der hypothetische Urmensch wohl ein vollentwickeltes
Parietalorgan, d. i. eine auf der Schädeldecke vollentwickelte
augenartige Öffnung, die jenen ältesten Landbewohnern
durchweg gemeinsam war und als ein bestimmtes, bisher nicht deutbares,
wenn auch sicher augenförmig ausgebildetes Sinnesorgan gelten
darf. Der Urmensch war wohl von jeher ein Säugetier.
Diese Säugetiernatur war aber habituell durch die soeben
aufgezählten Merkmale verhüllt. Nach der vorhin
erwähnten Tatsache, daß im Spätpaläozoikum sich
sogar unter den Reptilien deutliche Säugetiercharaktere bemerkbar
machen, obwohl ein Reptil niemals ein Säugetier gewesen sein kann,
ist anzunehmen, daß damals der Säugetierstamm als solcher
entstand, aber unter dem äußeren Gewand der
Reptilhaftigkeit; ebenso der Mensch als landtierartiges Wesen.
Im mesozoischen Zeitalter wird der Urmensch - er wird in vielen Stämmen und Einzelformen verschiedener Gattung gelebt haben - im allgemeinen sein Scheitelorgan durch Rückbildung langsam verloren und nun seine Säugetiernatur deutlicher enthüllt haben. In der Triaszeit finden wir die ersten als solche erkennbaren und in diesem Sinne echten Säugetierreste; sie gehören dem Stamm der Beuteltiere an, und nur solche findet man das ganze mesozoische Zeitalter hindurch in seltenen Resten. Dies war die damalige Stufe des Säugetierwesens, eben als Zeitsignatur, und daher wird auch der äußerlich schon säugetierhafter aussehende Mensch die anatomischen Eigenschaften des Beuteltieres geteilt haben, wie er im paläozoischen Zeitalter die des Reptils und Amphibiums trug. Im mesozoischen Zeitalter erkannten wir auch die Epoche, wo der aufrechte Gang der Landtiere angestrebt wurde; wir übertragen dies entsprechend auf den damaligen Menschenstamm. Es war auch die Zeit, wo die verwachsenere ursprünglichere Extremität der vollendeteren mit den spreizbaren Fingern und dem opponierbaren Daumen Platz machte; so wird auch der Mensch in dieser Hinsicht uns ähnlicher geworden sein. Das Säugetier hatte kein Scheitelorgan mehr; der mesozoische Mensch hatte es also mehr und mehr rückgebildet und dafür die vollere Entwicklung der Schädelkapsel, die vollere Entwicklung des Großhirns erreicht; er muß einen gewölbteren Schädel mit einer abgesetzten, weniger flachen Stirn bekommen haben.
Mit der Kreidezeit, wenigstens der letzten Hälfte der Kreidezeit, wo wir die letzte Herausentwicklung der mit der Alttertiärepoche fertig dastehenden fünffingerigen typischen Säugetierwelt schon jenseits des Beuteltierzustandes anzunehmen haben, wird dann auch der Säugetiermensch sich stark jenem Zustand genähert haben, der uns im Eiszeitmenschen fertig vor Augen tritt. Indessen ist der Eiszeitmensch als degenerierter Abkömmling des Spättertiärmenschen sehr stark pithekoid gestaltet, weil damals, wie schon gezeigt, der affenschaffende Zeitcharakter herrschte. Zuvor also muß der Mensch in seinen verschiedenen Spezialstämmen allerlei äußere Merkmale besessen haben, wie sie für die einzelnen Zeitstufen der Tertiärzeit gelten; so werden einzelnen seiner Stämme Fleischfresser- und Pflanzenfressermerkmale, ins Extrem entwickelt, teilweise nicht gefehlt haben. Schließlich endete der Menschenstamm unter Ausstoßung aller nicht zum Spätzeittypus gehörenden tierischen Charaktere in unserem heutigen Menschenstadium, das gewiß nicht einheitlich, sondern auf vielen Stammlinien wird erreicht worden sein und das nur deshalb anatomisch so einheitlich erscheint, weil eben jetzt die Zeitsignatur unseres Menschenstadiums herrscht. Dieses bevölkert heute die Erde, wie im paläozoischen Zeitalter wohl das scheiteläugige amphibienhafte, im mesozoischen Zeitalter das beuteltierhafte Menschenwesen.
Im mesozoischen Zeitalter wird der Urmensch - er wird in vielen Stämmen und Einzelformen verschiedener Gattung gelebt haben - im allgemeinen sein Scheitelorgan durch Rückbildung langsam verloren und nun seine Säugetiernatur deutlicher enthüllt haben. In der Triaszeit finden wir die ersten als solche erkennbaren und in diesem Sinne echten Säugetierreste; sie gehören dem Stamm der Beuteltiere an, und nur solche findet man das ganze mesozoische Zeitalter hindurch in seltenen Resten. Dies war die damalige Stufe des Säugetierwesens, eben als Zeitsignatur, und daher wird auch der äußerlich schon säugetierhafter aussehende Mensch die anatomischen Eigenschaften des Beuteltieres geteilt haben, wie er im paläozoischen Zeitalter die des Reptils und Amphibiums trug. Im mesozoischen Zeitalter erkannten wir auch die Epoche, wo der aufrechte Gang der Landtiere angestrebt wurde; wir übertragen dies entsprechend auf den damaligen Menschenstamm. Es war auch die Zeit, wo die verwachsenere ursprünglichere Extremität der vollendeteren mit den spreizbaren Fingern und dem opponierbaren Daumen Platz machte; so wird auch der Mensch in dieser Hinsicht uns ähnlicher geworden sein. Das Säugetier hatte kein Scheitelorgan mehr; der mesozoische Mensch hatte es also mehr und mehr rückgebildet und dafür die vollere Entwicklung der Schädelkapsel, die vollere Entwicklung des Großhirns erreicht; er muß einen gewölbteren Schädel mit einer abgesetzten, weniger flachen Stirn bekommen haben.
Mit der Kreidezeit, wenigstens der letzten Hälfte der Kreidezeit, wo wir die letzte Herausentwicklung der mit der Alttertiärepoche fertig dastehenden fünffingerigen typischen Säugetierwelt schon jenseits des Beuteltierzustandes anzunehmen haben, wird dann auch der Säugetiermensch sich stark jenem Zustand genähert haben, der uns im Eiszeitmenschen fertig vor Augen tritt. Indessen ist der Eiszeitmensch als degenerierter Abkömmling des Spättertiärmenschen sehr stark pithekoid gestaltet, weil damals, wie schon gezeigt, der affenschaffende Zeitcharakter herrschte. Zuvor also muß der Mensch in seinen verschiedenen Spezialstämmen allerlei äußere Merkmale besessen haben, wie sie für die einzelnen Zeitstufen der Tertiärzeit gelten; so werden einzelnen seiner Stämme Fleischfresser- und Pflanzenfressermerkmale, ins Extrem entwickelt, teilweise nicht gefehlt haben. Schließlich endete der Menschenstamm unter Ausstoßung aller nicht zum Spätzeittypus gehörenden tierischen Charaktere in unserem heutigen Menschenstadium, das gewiß nicht einheitlich, sondern auf vielen Stammlinien wird erreicht worden sein und das nur deshalb anatomisch so einheitlich erscheint, weil eben jetzt die Zeitsignatur unseres Menschenstadiums herrscht. Dieses bevölkert heute die Erde, wie im paläozoischen Zeitalter wohl das scheiteläugige amphibienhafte, im mesozoischen Zeitalter das beuteltierhafte Menschenwesen.
Der Mensch und das
Wirbeltier überhaupt, mit Einschuß auch der ältesten
Fische, hat kaum etwas in seinem Organismus, was erlaubte, ihn mit dem
ganzen Stamm der höheren Tiere an niedere Formen, also etwa Krebse
oder Würmer anzureihen. Ein kleines, jetzt lebendes
Fischchen, Branchiostoma,
äußerlich wurmförmig, aber vielleicht keine
ursprüngliche, sondern eine durch Rückbildung scheinbar
primitiv gewordene Gattung, von der man das Bild des Urahnen der
Wirbeltiere abnehmen wollte, unterscheidet sich ebenso prinzipiell von
allen Krebsen oder Würmern durch die grundlegend andersartige
Achsenlage des Gesamtkörpers; es hat einen Rückenstrang, ein
Rückenmark, während alle jene niederen Tiere ein Bauchmark
als zentralen Nervenstrang haben. Die Kluft scheint damit
unüberbrückbar. Aber doch gibt es wenigstens einen
Anhaltspunkt, daß im menschlichen Organismus, wie in dem der
späteren Wirbeltiere, eine Eigentümlichkeit übrig
geblieben sein könnte, welche tatsächlich auf ein noch
älteres Stadium als das des Uramphibiums oder des Fisches deutet
und in die Welt jener niederen Tiere unmittelbar
zurückweist. Das merkwürdige Organ des Scheitelauges,
das geradezu ein Symbol des Menschenwerdens sein kann, weist uns auch
hier wie auf eine älteste Spur seiner Herkunft hinab. Beim
Menschen zeigt sich, wie schon betont, das uralte Organ in zwiefacher
Reduktion. Einmal ist die Zirbeldrüse selbst schon ein
rückgebildetes Auge; sie hat aber noch einen seitlichen Begleiter,
der den Eindruck einer noch weiter fortgeschrittenen Rückbildung
macht und uns die Annahme nahelegt, daß ehedem das
vollentwickelte Parietalorgan doppelt gewesen sein muß. Da
sich aber sein noch weniger rückgebildeter Teil vergleichend
anatomisch als ein altes Auge erweist, so müßte das ehedem
vollentwickelte Organ zwei unmittelbar nebeneinander stehende Augen
dargestellt haben.
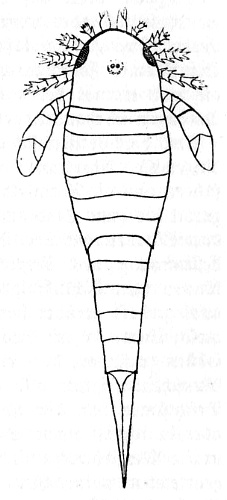
Fig. 10 (Bildquelle/-text:
"Urwelt, Sage und Menschheit" von Edgar Dacqué, 8. Aufl. 1938,
R.
Oldenbourg)
Krebsartiges Tier aus altpaläozoischer Zeit. Nordamerika. Mit zwei kleinen in der Mitte des Kopfes liegenden augenartigen Ozellen und großen,
nach vorne gerückten, randständigen Normalaugen. 1/2 nat. Gr. (Aus J. M. Clarke und R. Ruedemann, Eurypterida of New York. 1912.)
Krebsartiges Tier aus altpaläozoischer Zeit. Nordamerika. Mit zwei kleinen in der Mitte des Kopfes liegenden augenartigen Ozellen und großen,
nach vorne gerückten, randständigen Normalaugen. 1/2 nat. Gr. (Aus J. M. Clarke und R. Ruedemann, Eurypterida of New York. 1912.)
Nun finden wir
unter den alt- und mittelpaläozoischen Fischen und auch bei
krebsartigen Tieren ein derartiges vollentwickeltes doppeltes
Augenpaar, oben auf dem Schädeldach, obwohl daneben oder weiter
seitlich oder vorwärts noch zwei richtige Normalaugen liegen (Fig.
10). So läßt sich also hier vielleicht noch ein
schwacher Strahl einer Zeitsignatur, nicht einer Abstammung, auffangen,
die auch einmal bei des Menschenwesens frühesten körperlichen
Anfängen vorhanden gewesen sein könnte.
Doch dort verliert
sich der vergleichend anatomische Befund ins Ungewisse. Aber
innerhalb des übrigen Wirbeltierstammes selbst führen uns die
Formbildungen des Menschen in der ganzen Reihe zurück bis zum
Stadium des uramphibischen Formdaseins. Und damit haben wir den
Boden gewonnen für eine neue Altersbestimmung des Menschentypus,
des Menschenstammes, und können nun Schlußfolgerungen
ziehen, an die man bisher nur analogiehaft gerührt hat. Wir
dürfen erwarten, schon im Altmesozoikum, ja im
Spätpaläozoikum den Menschenstamm als solchen zu finden. d.
h. ein Wesen, das sich entelechisch durch seine Menschenhaftigkeit,
also auch durch gewisse seelische und geistige Besitztümer von der
übrigen Tierwelt unterschied. Wenn dies aber in irgend einem
Sinne zutrifft, so ist es auch möglich, daß uralte
Menschheitszustände und Erlebnisse aus einer ganz anders gearteten
natürlichen und seelischen Welt noch in späteren Mythen und
Sagen durchklingen.
Prof. Dr. Edgar Dacqué
Textanmerkungen/Spezialnachweise:
(1) Über den von der
bisherigen Deszendenztheorie in seinem Wesen falsch aufgefaßten
Begriff des "Stammbaumes" siehe: Kurt Lewin: "Die
Verwandtschaftsbegriffe in Biologie und Physik und die Darstellung
vollständiger Stammbäume". (In: Abhandlungen z.
theoret. Biologie v. J. Schaxel, Heft 5. Berlin 1920.) Ad. Raef,
Idealistische Morphologie u. Phylogenetik. Jena 1919.
Eine methodische Darstellung der Abstammungslehre auf historisch-kritischer Grundlage gibt S. Tschulok, Deszendenzlehre (Entwicklungslehre). Jena 1922.
Eine methodische Darstellung der Abstammungslehre auf historisch-kritischer Grundlage gibt S. Tschulok, Deszendenzlehre (Entwicklungslehre). Jena 1922.
2) L. Frobenius , Das
unbekannte Afrika. Aufhellung der Schicksale eines Erdteils.
München 1923, S. 23, 26.
Auf den hypothetischen, unzugehauenen Steinwerkzeugen aus der belgischen Tertiärformation, den Eolithen, baute schon früher Rutot einen entsprechenden Gedanken auf. Es sei anzunehmen, daß der Mensch ursprünglich nicht selbstgeschaffene, und seien es auch nur roh zugehauene Steinwerkzeuge benützt habe, sondern daß er schlechthin zu einem Gegenstand, also etwa zu einem geeigneten, von der Natur geformten Stein gegriffen habe. Dieser Zustand seiner Primitivkultur habe gewiß länger gedauert als der spätere steinzeitliche, und Rutot verlegt ihn nach den Eolithenfunden in die Jungtertiärzeit (Miozän). (M. Rutot, L 'etat actuel de la question de l 'antiquité de l 'homme. Bulletin Soc. belge de Géologie, Paléontologie et Hydrologie, Bruxelles 1903. T. XVII. S. 425-438). Allen derartigen Vorstellungen und Postulaten haftet aber der alte Fehler an, den auch die Abstammungslehre der Tiere und Pflanzen immer wieder macht, daß man nach einer formalen Primitivität sucht, die weder in der Natur, noch im Menschenleben als zeitlicher Urzustand bestanden hat und die, wenn irgendwo, dann ebenso gut auch später erst erscheinen kann und gar nicht der historische oder naturhistorische Ausgangspunkt für etwas unserem Auge oder unserem schematisierenden Abstraktionsvermögen kompliziert Erscheinendes zu sein braucht.
Auf den hypothetischen, unzugehauenen Steinwerkzeugen aus der belgischen Tertiärformation, den Eolithen, baute schon früher Rutot einen entsprechenden Gedanken auf. Es sei anzunehmen, daß der Mensch ursprünglich nicht selbstgeschaffene, und seien es auch nur roh zugehauene Steinwerkzeuge benützt habe, sondern daß er schlechthin zu einem Gegenstand, also etwa zu einem geeigneten, von der Natur geformten Stein gegriffen habe. Dieser Zustand seiner Primitivkultur habe gewiß länger gedauert als der spätere steinzeitliche, und Rutot verlegt ihn nach den Eolithenfunden in die Jungtertiärzeit (Miozän). (M. Rutot, L 'etat actuel de la question de l 'antiquité de l 'homme. Bulletin Soc. belge de Géologie, Paléontologie et Hydrologie, Bruxelles 1903. T. XVII. S. 425-438). Allen derartigen Vorstellungen und Postulaten haftet aber der alte Fehler an, den auch die Abstammungslehre der Tiere und Pflanzen immer wieder macht, daß man nach einer formalen Primitivität sucht, die weder in der Natur, noch im Menschenleben als zeitlicher Urzustand bestanden hat und die, wenn irgendwo, dann ebenso gut auch später erst erscheinen kann und gar nicht der historische oder naturhistorische Ausgangspunkt für etwas unserem Auge oder unserem schematisierenden Abstraktionsvermögen kompliziert Erscheinendes zu sein braucht.
3) G. Schwalbe, Die
Abstammung des Menschen und die ältesten Menschenformen. In:
Die Kultur der Gegenwart. III. Teil, Anthropologie. Leipzig
u. Berlin 1923, S. 316 u. 336 (Vgl. Anm. 5)
4) H. Klaatsch, Werdegang
der Menschheit. S. 17; S. 45ff. Ferner: Die Stellung des
Menschen im Naturganzen. XII. Vortrag in dem Sammelwerk: "Die
Abstammungslehre". Jena 1911. S. 332ff.
5) Hier und im Text kann
nicht der Beweis geführt werden, welche Theorie den Fossilfunden
und sonstigen Tatsachen am angemessensten erscheint. Es seien nur
kurz einige von Klaatsch gewählte Argumente erwähnt, aus
denen sich sein Urteil über die Stellung des Menschen zu den
Primaten ergibt. Für das Weitere muß auf die zitierten
Arbeiten und auf die gegnerische Stellung besonders von Schwalbe, den
wir nur als Hauptvertreter der ganzen Gegenrichtung nennen, verwiesen
werden.
Vorausnehmend sei zum Argument der Hand bemerkt, daß Schwalbe wohl nicht das Entscheidende in Klaatschs Argumentierung trifft, wenn er sagt, Klaatsch habe bei seiner Auffassung der Menschenhand und ihrer Verwertung als Beweisstück für ein hohes erdgeschichtliches Alter des Menschenstammes in erster Linie ihre Ausbildung als Greif- und Kletterorgan im Auge gehabt, wie sie sich im ältesten Tertiärabschnitt (Eozän) bei Halbaffen und früher noch bei Beuteltieren finde; er vergesse, daß bei jenen ältesten Säugetierformen die Hand stets ein Lokomotionsorgan, also gewissermaßen ein Fuß sei.
Klaatsch hat nicht behauptet, daß die Menschenhand wirklich von diesen alten Formen herkomme, und daß diese Formen die Stammeltern des Menschen seien, sondern er sagt, worin ich ihm unbedingt zustimme, daß der Primatenstamm, insbesondere der mit Menschenhandcharakter, schon mindestens im ältesten Tertiärzeitalter neben jenen fünffingerigen Extremitätenträgern bestanden haben muß, eben weil seine Hand sich prinzipiell von jener Extremität unterscheidet und jene Formen ohnehin in anderen Richtungen schon ganz und gar spezialisierte Typen waren. Da der Primatenstamm - so ist der Sinnzusammenhang in Klaatschs Beweisketten - aber nicht den Menschen entließ, sondern selbst eher der Abkömmling oder ein älterer, neben dem Menschenweg herlaufender Seitenzweig ist, so müsse auch aus diesem Grund der Mensch mindestens alttertiär, wenn nicht schon mesozoisch sein. Aus welchen Wahrscheinlichkeitsgründen das letztere möglich erscheint, ist im Text teils mit Klaatschs, teils mit eigenen Argumenten ja ausführlich dargelegt.
Vorausnehmend sei zum Argument der Hand bemerkt, daß Schwalbe wohl nicht das Entscheidende in Klaatschs Argumentierung trifft, wenn er sagt, Klaatsch habe bei seiner Auffassung der Menschenhand und ihrer Verwertung als Beweisstück für ein hohes erdgeschichtliches Alter des Menschenstammes in erster Linie ihre Ausbildung als Greif- und Kletterorgan im Auge gehabt, wie sie sich im ältesten Tertiärabschnitt (Eozän) bei Halbaffen und früher noch bei Beuteltieren finde; er vergesse, daß bei jenen ältesten Säugetierformen die Hand stets ein Lokomotionsorgan, also gewissermaßen ein Fuß sei.
Klaatsch hat nicht behauptet, daß die Menschenhand wirklich von diesen alten Formen herkomme, und daß diese Formen die Stammeltern des Menschen seien, sondern er sagt, worin ich ihm unbedingt zustimme, daß der Primatenstamm, insbesondere der mit Menschenhandcharakter, schon mindestens im ältesten Tertiärzeitalter neben jenen fünffingerigen Extremitätenträgern bestanden haben muß, eben weil seine Hand sich prinzipiell von jener Extremität unterscheidet und jene Formen ohnehin in anderen Richtungen schon ganz und gar spezialisierte Typen waren. Da der Primatenstamm - so ist der Sinnzusammenhang in Klaatschs Beweisketten - aber nicht den Menschen entließ, sondern selbst eher der Abkömmling oder ein älterer, neben dem Menschenweg herlaufender Seitenzweig ist, so müsse auch aus diesem Grund der Mensch mindestens alttertiär, wenn nicht schon mesozoisch sein. Aus welchen Wahrscheinlichkeitsgründen das letztere möglich erscheint, ist im Text teils mit Klaatschs, teils mit eigenen Argumenten ja ausführlich dargelegt.
Klaatsch Beweise
für die Selbständigkeit des Menschen
gegenüber dem Primatenstamm späterer Zeit, wie auch
gegenüber den übrigen tertiärzeitlichen Säugetieren
sind u. a. folgende:
1. Die Ursprünglichkeit der Menschenhand besteht in der formalen Annäherung an die fünffingerige Landextremität paläozoischer Amphibien. In ihrer Embryonalentwicklung aber zeigt sie eine rundliche ruderblattartige Form, vergleichbar jener von noch älteren paläozoischen Fischen. Ihre Flossenartigkeit wird noch besonders betont durch die Armabschnitte, die kurz sind und erst mit der Ausbildung zur eigentlichen Hand sich strecken. Die Scheidung von Ober- und Unterarm ist der Ausdruck für den ehemaligen Übergang vom Wasser- zum Landleben. Die Menschenhand läßt in dem Mechanismus ihrer Drehbewegungen noch die alten Ruderbewegungen erkennen, nämlich in der Rollung nach außen und innen, dem die Grundanordnung der Muskeln am Vorderarm noch entspricht.
2. Die Affen haben das auszeichnende Merkmal der Menschenhand, die Opponierbarkeit des Daumens, schon rückgebildet oder ganz verloren. Hierin ist der Mensch primitiver geblieben, kann also nicht von ihnen abgeleitet werden. Die Halbaffen allein haben die menschenartige Hand vollkommen bewahrt. Aber die Halbaffen sind nicht die unmittelbaren Ahnen der höheren Affen, auch nicht des Menschen, weil sie trotz Bewahrung einiger sehr ursprünglicher Merkmale doch einseitig differenziert sind. Wenn sie also mit den Primaten einschließlich des Menschen in wurzelechtem Zusammenhang stünden, dann könnte die alte Nahtstelle nur im mesozoischen Zeitalter liegen, weil sie schon in der Alttertiärzeit stark spezialisiert, d. i. vom Menschentypus stark verschieden erscheinen. So sind die Halbaffen eine Gruppe, die sich auf eigener Entwicklungsbahn von einem vielleicht mit den Primaten und dem Menschen gemeinsamen sehr alten Formstadium wegentwickelte und selbständig wie der Mensch den opponierbaren Daumen erworben haben kann.
3. Die Menschenhand hat zwei Elemente in sich: das des Greiffußes und das des Gehfußes. Sie hat an der Kleinfingerseite einen Muskelballen, der nicht zur Fingerbewegung dient, sondern ein Polster zum Schutz der Nerven und Gefäße gegen Pressung ist. Dieses Polster und der muskulöse Daumenballen bieten daher eine Sohlenfläche zu einer Art Laufunterstützung der Füße durch die Hände dar, wenn auch in sekundärer Entwicklung. Aber gerade, daß dies sekundär entwickelt ist und nicht primär, beweist, daß das vorhergehende ursprünglichere Stadium der Menschenhand eben nicht der einfache Lauffuß wie bei allen tertiärzeitlichen Säugetieren gewesen war.
4. Daß am Menschenfuß der Charakter einer Greifhand, also nicht einer einfachen Laufextremität wie bei den übrigen Säugetieren, ursprünglich da war, geht aus der Embryonalentwicklung hervor, die den Fuß nach Art der Hand zuerst noch gebaut zeigt, wobei die große Zehe wie ein Daumen absteht. Der jetzige Menschenfuß ist somit rückgebildet aus einem Kletterfuß, nicht aus einem Säugetierschreitfuß. Der Menschenaffentypus aber hat mit seinem Fuß einen so einseitig vorgeschrittenen Zustand schon erreicht, daß er als Seitenzweig des Urmenschentypus, nicht aber als Stammvater des Menschen darin erscheint. Auch die Anordnung der Blutgefäße und Nerven im Menschenfuß ist derart, daß der Raum zwischen 1. und 2. Zehe besonders betont ist, obwohl er äußerlich im fertigen Menschenfuß nicht mehr zum Ausdruck kommt.
1. Die Ursprünglichkeit der Menschenhand besteht in der formalen Annäherung an die fünffingerige Landextremität paläozoischer Amphibien. In ihrer Embryonalentwicklung aber zeigt sie eine rundliche ruderblattartige Form, vergleichbar jener von noch älteren paläozoischen Fischen. Ihre Flossenartigkeit wird noch besonders betont durch die Armabschnitte, die kurz sind und erst mit der Ausbildung zur eigentlichen Hand sich strecken. Die Scheidung von Ober- und Unterarm ist der Ausdruck für den ehemaligen Übergang vom Wasser- zum Landleben. Die Menschenhand läßt in dem Mechanismus ihrer Drehbewegungen noch die alten Ruderbewegungen erkennen, nämlich in der Rollung nach außen und innen, dem die Grundanordnung der Muskeln am Vorderarm noch entspricht.
2. Die Affen haben das auszeichnende Merkmal der Menschenhand, die Opponierbarkeit des Daumens, schon rückgebildet oder ganz verloren. Hierin ist der Mensch primitiver geblieben, kann also nicht von ihnen abgeleitet werden. Die Halbaffen allein haben die menschenartige Hand vollkommen bewahrt. Aber die Halbaffen sind nicht die unmittelbaren Ahnen der höheren Affen, auch nicht des Menschen, weil sie trotz Bewahrung einiger sehr ursprünglicher Merkmale doch einseitig differenziert sind. Wenn sie also mit den Primaten einschließlich des Menschen in wurzelechtem Zusammenhang stünden, dann könnte die alte Nahtstelle nur im mesozoischen Zeitalter liegen, weil sie schon in der Alttertiärzeit stark spezialisiert, d. i. vom Menschentypus stark verschieden erscheinen. So sind die Halbaffen eine Gruppe, die sich auf eigener Entwicklungsbahn von einem vielleicht mit den Primaten und dem Menschen gemeinsamen sehr alten Formstadium wegentwickelte und selbständig wie der Mensch den opponierbaren Daumen erworben haben kann.
3. Die Menschenhand hat zwei Elemente in sich: das des Greiffußes und das des Gehfußes. Sie hat an der Kleinfingerseite einen Muskelballen, der nicht zur Fingerbewegung dient, sondern ein Polster zum Schutz der Nerven und Gefäße gegen Pressung ist. Dieses Polster und der muskulöse Daumenballen bieten daher eine Sohlenfläche zu einer Art Laufunterstützung der Füße durch die Hände dar, wenn auch in sekundärer Entwicklung. Aber gerade, daß dies sekundär entwickelt ist und nicht primär, beweist, daß das vorhergehende ursprünglichere Stadium der Menschenhand eben nicht der einfache Lauffuß wie bei allen tertiärzeitlichen Säugetieren gewesen war.
4. Daß am Menschenfuß der Charakter einer Greifhand, also nicht einer einfachen Laufextremität wie bei den übrigen Säugetieren, ursprünglich da war, geht aus der Embryonalentwicklung hervor, die den Fuß nach Art der Hand zuerst noch gebaut zeigt, wobei die große Zehe wie ein Daumen absteht. Der jetzige Menschenfuß ist somit rückgebildet aus einem Kletterfuß, nicht aus einem Säugetierschreitfuß. Der Menschenaffentypus aber hat mit seinem Fuß einen so einseitig vorgeschrittenen Zustand schon erreicht, daß er als Seitenzweig des Urmenschentypus, nicht aber als Stammvater des Menschen darin erscheint. Auch die Anordnung der Blutgefäße und Nerven im Menschenfuß ist derart, daß der Raum zwischen 1. und 2. Zehe besonders betont ist, obwohl er äußerlich im fertigen Menschenfuß nicht mehr zum Ausdruck kommt.
Zu entsprechenden
Ergebnissen führt die Betrachtung des Gebisses:
1. Affen- und Menschengebiß sind durch ihre ursprüngliche Lückenlosigkeit, die keinem jungtertiärzeitlichen Säugetier mehr zukommt, ausgezeichnet. Aber auch die in den Menschenstammbaum eingereihten Halbaffen haben in der Alttertiärzeit schon zwischen Eck- und Backenzähnen die Lücke. Nur bei einer Form (Necrolemur) aus der Alttertiärzeit erscheint die volle primatenartige Zahnzahl. Aber ihn deshalb in die Stammreihe der Primaten und des Menschen einzurücken, verbietet der starke Eckzahn, der zwar ein pithekoides, aber kein menschliches Merkmal ist, und außerdem hat er statt der Lücke ein an Zahnzahl geringeres Gebiß, ist somit hierin schon einseitig entwickelt.
2. Die jetzigen Menschenaffen haben, zum großen Unterschied auch vom Diluvialmenschen, stark entwickelte Eckzähne (Fig. 6). Würde der Mensch von Trägern eines solchen starken Eckzahnes abstammen, so müßte der Entwicklungslauf in einer Reduktion der Eckzahnstärke bestehen, mithin müßten die ältesten gemeinsamen Stammformen von Mensch und Primaten erst recht starke Eckzähne gehabt haben; auch der diluviale Mensch müßte als vermutlicher Vorfahre des Jetztzeitmenschen noch stärkere Eckzähne als dieser besessen haben, was nicht der Fall ist, obwohl der Schädel durch seine etwas niedrigere Wölbung und seine Augenwülste affenähnlicher war. Da aber auch die Halbaffen wegen ihrer sonstigen Differenzierung in Schädel und Gebiß nicht Stammeltern des Menschen sein können, so ist für den Menschenstamm die Bahn außerhalb der Primaten und der Halbaffen zunächst bis in die Alttertiärstufe hinunter frei.
1. Affen- und Menschengebiß sind durch ihre ursprüngliche Lückenlosigkeit, die keinem jungtertiärzeitlichen Säugetier mehr zukommt, ausgezeichnet. Aber auch die in den Menschenstammbaum eingereihten Halbaffen haben in der Alttertiärzeit schon zwischen Eck- und Backenzähnen die Lücke. Nur bei einer Form (Necrolemur) aus der Alttertiärzeit erscheint die volle primatenartige Zahnzahl. Aber ihn deshalb in die Stammreihe der Primaten und des Menschen einzurücken, verbietet der starke Eckzahn, der zwar ein pithekoides, aber kein menschliches Merkmal ist, und außerdem hat er statt der Lücke ein an Zahnzahl geringeres Gebiß, ist somit hierin schon einseitig entwickelt.
2. Die jetzigen Menschenaffen haben, zum großen Unterschied auch vom Diluvialmenschen, stark entwickelte Eckzähne (Fig. 6). Würde der Mensch von Trägern eines solchen starken Eckzahnes abstammen, so müßte der Entwicklungslauf in einer Reduktion der Eckzahnstärke bestehen, mithin müßten die ältesten gemeinsamen Stammformen von Mensch und Primaten erst recht starke Eckzähne gehabt haben; auch der diluviale Mensch müßte als vermutlicher Vorfahre des Jetztzeitmenschen noch stärkere Eckzähne als dieser besessen haben, was nicht der Fall ist, obwohl der Schädel durch seine etwas niedrigere Wölbung und seine Augenwülste affenähnlicher war. Da aber auch die Halbaffen wegen ihrer sonstigen Differenzierung in Schädel und Gebiß nicht Stammeltern des Menschen sein können, so ist für den Menschenstamm die Bahn außerhalb der Primaten und der Halbaffen zunächst bis in die Alttertiärstufe hinunter frei.
Auch die Schädelbildung
liefert noch einige Anhaltspunkte, die kurz erwähnt seien:
1. Beim menschlichen Embryo verlagern sich die Augen von der Seite nach vorne. Der fertige Mensch ist in der nach vorne gerichteten Stellung seiner Augen das vollkommenste Säugetier. Diese Vollkommenheit ist aber nicht gleich der extremsten Entwicklung in dieser Richtung. Denn die Menschenaffen sind darin übertrieben spezialisiert, über den Menschenzustand noch hinaus. Mit jener Augenverlagerung nach vorne wird die ursprüngliche säugetierhafte Nasenregion verschmälert. Bei den Affen ist diese noch schmäler geworden als beim Menschen, dessen Augenhöhlen noch seitlicher liegen, wenn sie auch im Gegensatz zu den übrigen Säugetieren sehr nach vorne gerückt sind. Die Affen können also auch hierin nicht das Vorstadium zum Menschen sein.
2. Mit der Augenverlagerung hängt auch die Schädelwölbung zusammen. Durch die Verdrängung des Geruchsorgans bei der Vorwärtsverlagerung der Augen, durch die damit eingetretene Erweiterung der Gesichtseindrücke konnte sich die Großhirnhemisphäre beim Menschen so erweitern, daß er darin alle anderen Tiere übertraf. Solange das Schädeldach noch flach war oder nach hinten anstieg, war die vordere Augenregion von der hinteren Schädelregion abgegrenzt. Als sich das Großhirn und damit das Schädeldach durch seine Wölbung über die vordere Region emporhob, blieben als betonter Rest jene Augenwülste übrig, die den Diluvialmensch und den Australier noch auszeichnen. Beim Gorilla setzt sich die besonders starke Kiefermuskulatur an die Augenwülste an und verstärkt sie noch. In dem entwicklungsgeschichtlichen Augenblick, wo die Schädelwölbung in dem beschriebenen Zusammenhang einsetzte, war der Divergenzpunkt einerseits zum heutigen Menschen, andererseits zu den Menschenaffen erreicht. Von da ab mußte theoretisch einerseits der Mensch mit der hochgewölbten Stirn, andererseits der immer menschenaffenartiger werdende Gorillastamm sich abzweigen, dessen Extrem schließlich der Gorillaschädel mit dem Knochenkamm über das Schädeldach herüber wurde. Das Junge des Gorilla hat noch einen sehr menschenähnlichen Schädel im Gegensatz zum erwachsenen Tier. Nachdem der Gorillastamm sich abgezweigt hatte, ging der noch nicht vollendete Menschenstamm seinen Weg weiter zum vollendeten Großhirn mit dem gewölbten Schädel; auf einem dabei erreichten höheren Entwicklungsstadium stellte sich noch einmal eine Spaltung ein, die den übrigen Menschenaffenkomplex schuf. Und damit erst war der vollendete quartärzeitliche Vollmensch da.
1. Beim menschlichen Embryo verlagern sich die Augen von der Seite nach vorne. Der fertige Mensch ist in der nach vorne gerichteten Stellung seiner Augen das vollkommenste Säugetier. Diese Vollkommenheit ist aber nicht gleich der extremsten Entwicklung in dieser Richtung. Denn die Menschenaffen sind darin übertrieben spezialisiert, über den Menschenzustand noch hinaus. Mit jener Augenverlagerung nach vorne wird die ursprüngliche säugetierhafte Nasenregion verschmälert. Bei den Affen ist diese noch schmäler geworden als beim Menschen, dessen Augenhöhlen noch seitlicher liegen, wenn sie auch im Gegensatz zu den übrigen Säugetieren sehr nach vorne gerückt sind. Die Affen können also auch hierin nicht das Vorstadium zum Menschen sein.
2. Mit der Augenverlagerung hängt auch die Schädelwölbung zusammen. Durch die Verdrängung des Geruchsorgans bei der Vorwärtsverlagerung der Augen, durch die damit eingetretene Erweiterung der Gesichtseindrücke konnte sich die Großhirnhemisphäre beim Menschen so erweitern, daß er darin alle anderen Tiere übertraf. Solange das Schädeldach noch flach war oder nach hinten anstieg, war die vordere Augenregion von der hinteren Schädelregion abgegrenzt. Als sich das Großhirn und damit das Schädeldach durch seine Wölbung über die vordere Region emporhob, blieben als betonter Rest jene Augenwülste übrig, die den Diluvialmensch und den Australier noch auszeichnen. Beim Gorilla setzt sich die besonders starke Kiefermuskulatur an die Augenwülste an und verstärkt sie noch. In dem entwicklungsgeschichtlichen Augenblick, wo die Schädelwölbung in dem beschriebenen Zusammenhang einsetzte, war der Divergenzpunkt einerseits zum heutigen Menschen, andererseits zu den Menschenaffen erreicht. Von da ab mußte theoretisch einerseits der Mensch mit der hochgewölbten Stirn, andererseits der immer menschenaffenartiger werdende Gorillastamm sich abzweigen, dessen Extrem schließlich der Gorillaschädel mit dem Knochenkamm über das Schädeldach herüber wurde. Das Junge des Gorilla hat noch einen sehr menschenähnlichen Schädel im Gegensatz zum erwachsenen Tier. Nachdem der Gorillastamm sich abgezweigt hatte, ging der noch nicht vollendete Menschenstamm seinen Weg weiter zum vollendeten Großhirn mit dem gewölbten Schädel; auf einem dabei erreichten höheren Entwicklungsstadium stellte sich noch einmal eine Spaltung ein, die den übrigen Menschenaffenkomplex schuf. Und damit erst war der vollendete quartärzeitliche Vollmensch da.
Fr. Weidenreich, Der
Menschenfuß (Zeitschr. für Morphologie und Anthropologie.
Bd. 22. Stuttgart 1921, S. 51-282) kommt bei der anatomischen und
statischen Durcharbeitung des Menschenfußes dazu, den Menschen
von Formen mit fünffingerigem Kletterfuß theoretisch
abzuleiten, worin er sich als primitivste und beweglichste Form
gegenüber allen Säugetieren, außer den kletternden
Primaten und Beuteltieren, erweist. Diesen gegenüber ist der
Menschenfuß aber durchaus einseitig spezialisiert. Der
Fuß des Jetztweltmenschen zeigt trotz bemerkenswerter primitiver
Merkmale einzelner Rassen keine direkten Beziehungen zu einer
bestimmten Primatengruppe. Der Hominidenahn muß schon von
vornherein lange untere Extremitäten besessen haben, als er so
die terrestrische Lebensweise aufnahm. Die anthropomorphen Affen
scheiden aus. Es muß einmal eine Primatenform bestanden
haben, die wie die Beuteltiere eine besondere Hautverbindung zwischen
zweier und dritter Zehe (Zygodaktylie) neben einer allgemeinen
Schwimmhautbildung besaß. Diese ging teilweise verloren,
wurde aber im Hylobatiden- und Hominidenstamm bewahrt und beweist auch
die stammesgeschichtliche Selbständigkeit des Sprosses, der zum
Menschen führte. Wo er abzweigte, ist schwer zu sagen.
Weidenreich nimmt mit
Schwalbe an, daß der Sproß "in seiner Selbständigkeit
sehr weit herabreicht".
6) M.
Schlosser (Beiträge zur
Kenntnis der oligozänen Landsäugetiere aus dem Fayûm,
Ägypten. In: Beiträge z. Paläontologie und
Geologie Österr.-Ungarns und des Orients.
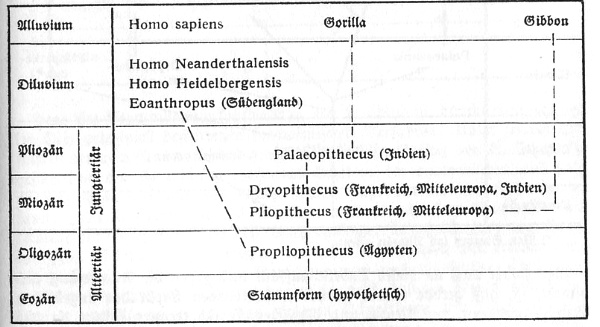
Bd. 24. Wien-Leipzig
1911, S. 55ff.) verfolgt den "Stammbaum" der
Simiiden bis zu diesem Propliopithecus zurück, und danach
ergäbe sich umstehende Ahnenreihe, zu der die einzelnen Gattungen
in kurzen Definitionen und teilweise mit Abbildung ihrer Reste in dem
Lehrbuch von K. A. v. Zittel (Grundzüge der
Paläontologie. Bd. II. Wirbeltiere. 4.Aufl.
München-Berlin 1923. Säugetiere, bearb. von M.
Schlosser) aufgeführt sind.
Die Gattungen dieses Stammbaumes treten auf weite Länder verteilt auf und sind zum Teil nur auf Kieferreste (Fig. 6) oder bloß Einzelzähne gegründet, von denen es teilweise überhaupt zweifelhaft ist, ob sie zusammengehören (Eoanthropus). Mit diesem Stammbaum erklärt Schlosser den Tertiärmenschen und seine vermeintlichen Primitivwerkzeuge (Eolithen) für widerlegt, zumal auch der diluvale Primitivkiefer des Homo Heidelbergensis die Hypothese eines Tertiärmenschen überflüssig gemacht haben soll.
Die Gattungen dieses Stammbaumes treten auf weite Länder verteilt auf und sind zum Teil nur auf Kieferreste (Fig. 6) oder bloß Einzelzähne gegründet, von denen es teilweise überhaupt zweifelhaft ist, ob sie zusammengehören (Eoanthropus). Mit diesem Stammbaum erklärt Schlosser den Tertiärmenschen und seine vermeintlichen Primitivwerkzeuge (Eolithen) für widerlegt, zumal auch der diluvale Primitivkiefer des Homo Heidelbergensis die Hypothese eines Tertiärmenschen überflüssig gemacht haben soll.
W. K. Gregory, der mit
Schlosser an der Bedeutung des alttertiärzeitlichen
ägyptischen Propliopithecus als unmittelbarem
Menschenaffenahnen festhält, gibt folgenden, hier nur wesentlich
verkürzten Stammbaum (The origin and evolution of the human
dentition. Journ. of Dental Research. Vol. II S. 688. New Haven
1920. Ferner: Studies on the evolution of the Primates.
Bullet. Americ. Mus. Nat. Hist. New York 1916, S. 313ff.):
*) Von Gregory ins
Pliozän gesetzt.
Was sofort auch an
dieser Tabelle auffällt und gegen die Anordnung
einnimmt, ist, daß gerade wieder an der entscheidenden Stelle des
angeblichen Menschenwerdens, wo Mensch und Menschenaffe sich trennen
sollten, so daß man von einer nachweisbar späten Entstehung
des Menschen sprechen dürfte, eine imaginäre Stammform
angedeutet werden muß, ein Strichknoten statt eines wirklichen
Ahnherrn. Derart ist aber immer das Ergebnis der formalen
Deszendenzlehre, man mag es mit welchen Tieren nur immer, hohen oder
niederen, zu tun haben: immer dort, wo wir eine wirkliche "Urform",
einen wirklichen Stammvater haben sollten, müssen wir uns mit
Bindestrichen begnügen; die wirkliche Urform aber - in diesem
Falle Propliopithecus
- ist ein nicht einmal vollständiger Unterkiefer. Würde
zu diesem das ganze Skelett gefunden, so müßte es sich erst
noch zeigen, ob die Gattung nicht doch so einseitig entwickelt war,
daß sie wiederum aus diesem Stammbaum als Seitenzweig
auszuscheiden ist. Man bedenke auch, daß auf der
vorstehenden abgekürzten Tabelle sehr zugunsten dieses Stammbaumes
noch der Umstand spricht, daß hier einige Zeitstufen im
Miozän und Oligozän zusammengenommen sind, so daß in
Wirklichkeit Propliopithecus
noch weiter ohne sichere Zwischenglieder vom Menschen
abrückt. Entwerfen wir also mit den vorigen Mitteln den
Stammbaum ohne petitio principii,
dann sieht er wohl aus, wobei die Strichelung das Hypothetische
darstellt:
Jene Art
Stammbaumrekonstruktion, wie sie üblich ist, beruht eben
auf dem im Text prinzipiell bestrittenen formalistischen
Verfahren. Unter seinem Einfluß ist der auch für den
Nichtfachmann sehr lesbare Aufsatz von G. Schwalbe geschrieben, der als
zusammenfassendes Ergebnis der anatomischen und paläontologischen
Forschung des letzten halben Jahrhunderts angesehen werden kann: "Die
Abstammung des Menschen und die ältesten Menschenformen.
(Kultur der Gegenwart. Teil III. Anthropologie. Leipzig-Berlin 1923,
S. 223 bis 338). Schwalbe ist ein Gegner der Lehre von Klaatsch und erkennt weder die Herkunft der Affen aus dem Menschenstamm, noch die Existenz eines tertiär zeitlichen Menschen an. Wenn er darauf hinweist, daß der Mensch viele Eigenschaften enthält, die er mit den niederen Säugetieren teilt (Gaumenfalten-Jacobsohnsches Organ in der Nase, Übereinstimmung der Embryonalformen z. B. in den Kiemenspalten usw.) und ihn damit formal an die niederen Säugetiere anschließt, so wäre die gegebene Ausdrucksweise für diese Tatsachen nicht die, daß der Mensch "deshalb" von niederen Säugetieren "abstamme", sondern die, daß er niedere Säugetierstadien an sich hat; womit er ebensogut deren Ahne sein kann, wie auch beide einen gemeinsamen Ausgangspunkt haben können. Denn auch die niederen Säugetiere, soweit man zurückgeht, sind so einseitig entwickelt, daß nicht abzusehen ist, wo der Mensch mit seinen vielfach nicht nur sehr primitiven, sondern auch anders gearteten Eigentümlichkeiten später gegen Ende der Tertiärzeit angeknüpft werden könnte. Denn die von Schlosser gegebene Affenreihe - soweit wir nicht nur einzelne Zähne und Kieferreste miteinander vergleichen - ist schon, nach Klaatschs Darlegungen, mit einbegriffen in die ursprüngliche Primitivität insbesondere der Hand, so daß man den Menschen als solchen nicht erst kurz vor dem Diluvium aus jener entstehen lassen darf. Hierfür sei auf die schon angegebenen Arbeiten von Klaatsch selbst hingewiesen, deren Kritik dann in dem hier genannten Aufsatz von Schwalbe zu finden ist. Der wissenschaftliche Streit bekommt ein anderes Gesicht, wenn man die biologisch unhaltbare alte Stammbaumvorstellung als eine Fiktion und ein der Natur nicht entsprechendes Abstraktum aufgibt, sich das Wesen der Typentheorie und das Gesetz des Zeitcharakters klar macht und daraufhin die natürlich gegebenen Formen betrachtet, ohne sie in ein Schema zu drängen.
S. 223 bis 338). Schwalbe ist ein Gegner der Lehre von Klaatsch und erkennt weder die Herkunft der Affen aus dem Menschenstamm, noch die Existenz eines tertiär zeitlichen Menschen an. Wenn er darauf hinweist, daß der Mensch viele Eigenschaften enthält, die er mit den niederen Säugetieren teilt (Gaumenfalten-Jacobsohnsches Organ in der Nase, Übereinstimmung der Embryonalformen z. B. in den Kiemenspalten usw.) und ihn damit formal an die niederen Säugetiere anschließt, so wäre die gegebene Ausdrucksweise für diese Tatsachen nicht die, daß der Mensch "deshalb" von niederen Säugetieren "abstamme", sondern die, daß er niedere Säugetierstadien an sich hat; womit er ebensogut deren Ahne sein kann, wie auch beide einen gemeinsamen Ausgangspunkt haben können. Denn auch die niederen Säugetiere, soweit man zurückgeht, sind so einseitig entwickelt, daß nicht abzusehen ist, wo der Mensch mit seinen vielfach nicht nur sehr primitiven, sondern auch anders gearteten Eigentümlichkeiten später gegen Ende der Tertiärzeit angeknüpft werden könnte. Denn die von Schlosser gegebene Affenreihe - soweit wir nicht nur einzelne Zähne und Kieferreste miteinander vergleichen - ist schon, nach Klaatschs Darlegungen, mit einbegriffen in die ursprüngliche Primitivität insbesondere der Hand, so daß man den Menschen als solchen nicht erst kurz vor dem Diluvium aus jener entstehen lassen darf. Hierfür sei auf die schon angegebenen Arbeiten von Klaatsch selbst hingewiesen, deren Kritik dann in dem hier genannten Aufsatz von Schwalbe zu finden ist. Der wissenschaftliche Streit bekommt ein anderes Gesicht, wenn man die biologisch unhaltbare alte Stammbaumvorstellung als eine Fiktion und ein der Natur nicht entsprechendes Abstraktum aufgibt, sich das Wesen der Typentheorie und das Gesetz des Zeitcharakters klar macht und daraufhin die natürlich gegebenen Formen betrachtet, ohne sie in ein Schema zu drängen.
6a) A.
Adolff, Einige besondere
Bildungen an den Zähnen des Menschen und ihre Bedeutung für
seine Vorgeschichte. Anatom. Anzeiger v. Eggeling.
Bd. 58. Jena 1924, S. 497ff.
7) In diesem Sinne sagt
selbst Schwalbe, es gehe keinesfalls an, die Greifhand des Menschen auf
eine solche bei alttertiären Säugetieren
zurückzuführen. Beuteltiere kämen wegen ihrer
sonstigen Differenziertheit nicht in Betracht, es sei denn, daß
man an die anzunehmenden älteren gemeinsamen Ausgangsformen der
Plazentalier und Marsupialier anknüpfen wolle.
Man sieht, wie durch die sachliche Betrachtung auch dieser, ein höheres Alter des Menschen so von Grund aus ablehnende Forscher, ohne daß er es bemerkt, schon zu einem außerordentlich hohen Alter geführt wird.
Man sieht, wie durch die sachliche Betrachtung auch dieser, ein höheres Alter des Menschen so von Grund aus ablehnende Forscher, ohne daß er es bemerkt, schon zu einem außerordentlich hohen Alter geführt wird.
8) W. K. Gregory, R. W.
Miner, G.K. Noble, The Carpus of Eryops and the Structure of the
primitive Chiropterygium. Bullet. Americ. Mus. Nat. Hist.
New York 1923, Vol. 48, S. 279.
W. K. Gregory, Present status of the problem of the origin of the Tetrapoda etc. Annals New York Acad. Science. Vol. 26, 1915, S. 317-383.
W. K. Gregory, Present status of the problem of the origin of the Tetrapoda etc. Annals New York Acad. Science. Vol. 26, 1915, S. 317-383.
8a) W. Soergel. Die Fährten der Chirotheria. Eine paläobiologische Studie. Jena 1925.
9) Zu unterscheiden von
der gleichmäßigen Streckung der ganzen Extremität ist
deren sekundäre Verlängerung bei springenden Tieren wie
Vögeln und gewissen Säugetieren (Springhasen und -mäuse)
des känozoischen Zeitalters. Da werden nicht die Hinterbeine
als Ganzes gestreckt und der Fuß behält seine
Eigentümlichkeit bei, sondern die Fußwurzelknochen vor allem
werden stark verlängert und sonstwie noch modifiziert. Nur
bei einem gewissen Teil von Schrecksauriern aus der Jura- und
Kreidezeit tritt dieselbe Streckung der Fußwurzelknochen ein; das
sind jene Gattungen, welche etwas durchaus Vogelähnliches haben,
die wie ein Straußvogel aufrecht gingen und wegen ihrer hohlen
Knochen, sowie einigen anderen Skeletteigentümlichkeiten mit
Vögeln in stammesgeschichtlicher Beziehung stehen
können. Ihre zum Teil gewaltigen Dimensionen lassen in ihnen
die sagenhaften Drachen vermuten. Abermals davon zu unterscheiden
ist die Stelzbeinigkeit, welche nicht nur in den beiden
Hinterfüßen, sondern in allen vier Füßen
gleichartig erreicht ist durch Streckung und Verschmelzung einzelner
Fußwurzelknochen, wie bei Hirschen und Schafen. Besonders
wichtig zur Feststellung der Hochbeinigkeit als Zeitsignatur ist
ferner, daß auch die , jedenfalls den Typus des mesozoischen
Säugetieres ausschließlich ausmachenden Beuteltiere, soweit
sie aufrecht gehen und stehen, stets den in seiner Wurzel schon etwas
verlängerten Fuß, jedoch völlig auf dem Boden liegend
haben, so daß äußerlich durchaus der Habitus des
gewöhnlichen mesozoischen Landreptils mit den einfach gestreckten
Hinterbeinen gewahrt wird.
10) G. Steinmann, Der Ursprung des Menschen. Die Westmark. Köln-Mühlheim 1921, S. 457ff.
10a) M. Westenhöfer,
Über die Erhaltung von Vorfahrenmerkmalen beim Menschen usw.
Medizinische Klinik. Jahrg. 19. Berlin 1923, Nr. 37. -, Das
menschliche Kinn, seine Entstehung und anthropologische
Bedeutung. Archiv f. Frauenkunde u. Konstitutionsforschung.
Bd. X. Berlin 1924, S. 239-262.